DEZVOLTAREA embrionilor
Esența etapei de zdrobire. Despărțirea - aceasta este o serie de diviziuni mitotice succesive ale zigotului și ale altor blastomere, care se termină cu formarea unui embrion multicelular - blastula. Prima diviziune a zdrobirii începe după combinarea materialului ereditar al pronucleilor și formarea unei plăci metafazice comune. Celulele formate în timpul clivajului sunt numite blastomeri(din greaca. explozie- germen, germen). O caracteristică a diviziunilor mitotice de zdrobire este că, cu fiecare diviziune, celulele devin din ce în ce mai mici până când ajung la raportul dintre volumele nucleului și citoplasmei, care este obișnuit pentru celulele somatice. La un arici de mare, de exemplu, acest lucru necesită șase diviziuni, iar embrionul este format din 64 de celule. Între diviziunile succesive, creșterea celulară nu are loc, dar ADN-ul este neapărat sintetizat.
Toți precursorii ADN și enzimele necesare se acumulează în timpul oogenezei. Ca urmare, ciclurile mitotice sunt scurtate și diviziunile se succed mult mai repede decât în celulele somatice obișnuite. În primul rând, blastomerele sunt adiacente unul altuia, formând un grup de celule numite morula. Apoi se formează o cavitate între celule - blastocoel, umplut cu lichid. Celulele sunt împinse la periferie, formând peretele blastulei - blastoderm. Dimensiunea totală a embrionului până la sfârșitul clivajului în stadiul de blastula nu depășește dimensiunea zigotului.
Principalul rezultat al perioadei de zdrobire este transformarea zigotului în embrion multicelular cu un singur schimb.
Morfologia strivirii. De regulă, blastomerele sunt aranjate într-o ordine strictă unul față de celălalt și axa polară a oului. Ordinea sau metoda de zdrobire depinde de cantitatea, densitatea și natura distribuției gălbenușului în ou. Conform regulilor lui Sachs - Hertwig, nucleul celular tinde să fie situat în centrul citoplasmei fără gălbenuș, iar axul diviziunii celulare - în direcția cea mai mare a acestei zone.
În ouăle oligo- și mezolecitale, clivaj complet, sau holoblastic. Acest tip de strivire se găsește la lamprede, unii pești, toți amfibieni, precum și la marsupiale și mamiferele placentare. La zdrobire completă, planul primei diviziuni corespunde planului de simetrie bilaterală. Planul celei de-a doua diviziuni merge perpendicular pe planul primei. Ambele brazde ale primelor două diviziuni sunt meridiane, ᴛ.ᴇ. începe de la polul animal și se extinde la polul vegetativ. Celula ou este împărțită în patru blastomere mai mult sau mai puțin egale ca dimensiuni. Planul celei de-a treia diviziuni se desfășoară perpendicular pe primele două în direcția latitudinală. După aceea, în ouăle mesolecitale în stadiul de opt blastomere se manifestă zdrobirea neuniformă. La polul animal există patru blastomere mai mici - micrometre, pe vegetativ - patru mai mari - macromeri. Apoi diviziunea merge din nou în planurile meridiane și apoi din nou în latitudinal.
În ovocitele polilecitale ale peștilor osoși, reptilelor, păsărilor și, de asemenea, mamiferelor monotreme, clivajul parțial, sau meroblastic,ᴛ.ᴇ. acoperă doar citoplasma lipsită de gălbenuș. Este situat sub forma unui disc subțire la polul animalului, în legătură cu aceasta, acest tip de zdrobire se numește discoidal.
La caracterizarea tipului de zdrobire se ia în considerare și poziția relativă și rata de divizare a blastomerilor. Dacă blastomerele sunt aranjate în rânduri unul deasupra celuilalt de-a lungul razelor, se numește zdrobire radial. Este tipic pentru cordate și echinoderme. În natură, există și alte variante de aranjare spațială a blastomerelor în timpul zdrobirii, ceea ce determină astfel de tipuri de ele ca spirală la moluște, bilaterală la ascaris, anarhică la meduze.
S-a remarcat o relație între distribuția gălbenușului și gradul de sincronism în diviziunea blastomerelor animale și vegetative. În ouăle oligolecite ale echinodermelor, clivajul este aproape sincron; în celulele ouă mezocitetale, sincronia este perturbată după a treia diviziune, deoarece blastomerele vegetative se divid mai lent din cauza cantității mari de gălbenuș. În formele cu strivire parțială, diviziunile sunt asincrone de la bun început și blastomerele care ocupă o poziție centrală se divid mai repede.
Orez. 7.2. Clivaj în cordate cu diferite tipuri de ouă.
DAR - lanceta; B - broască; IN - pasăre; G - mamifer:
eu-două blastomere II- patru blastomeri, III- opt blastomeri, IV- morula, V- blastula;
1 - brazde de zdrobire, 2 -blastomere, 3- blastoderm, 4- blastoiel, 5- epiblast, 6- hipoblast, 7-embrioblast, 8- trofoblast; dimensiunile nucleelor din figură nu reflectă rapoartele reale de mărime
Orez. 7.2. Continuare
La sfârșitul zdrobirii, se formează o blastula. Tipul de blastula depinde de tipul de zdrobire și, prin urmare, de tipul de ou. Unele tipuri de zdrobire și blastula sunt prezentate în Fig. 7.2 și schema 7.1. Pentru o descriere mai detaliată a clivajului la mamifere și oameni, a se vedea Sec. 7.6.1.
Caracteristicile proceselor molecular-genetice și biochimice în timpul zdrobirii. După cum sa menționat mai sus, ciclurile mitotice în timpul perioadei de clivaj sunt mult scurtate, mai ales la început.
De exemplu, întregul ciclu de fisiune în ouăle de arici de mare durează 30-40 de minute, cu durata fazei S de doar 15 minute. Perioadele gi și 02 sunt practic absente, deoarece în citoplasma celulei ou s-a creat aprovizionarea necesară cu toate substanțele și, cu cât este mai mare, cu atât este mai mare. Înainte de fiecare diviziune are loc sinteza ADN-ului și a histonelor.
Rata cu care furculița de replicare se mișcă de-a lungul ADN-ului în timpul clivajului este normală. În același timp, există mai multe puncte de inițiere în ADN-ul blastomerilor decât în celulele somatice. Sinteza ADN-ului are loc în toți repliconii simultan, sincron. Din acest motiv, timpul de replicare a ADN-ului în nucleu coincide cu timpul de dublare a unuia, în plus, scurtat, replicon. S-a demonstrat că atunci când nucleul este îndepărtat din zigot, are loc clivajul, iar embrionul în dezvoltare ajunge aproape în stadiul de blastulă. Dezvoltarea ulterioară se oprește.
La începutul clivajului, alte tipuri de activitate nucleară, cum ar fi transcripția, sunt practic absente. În diferite tipuri de ouă, transcripția genelor și sinteza ARN încep în diferite etape. În cazurile în care există multe substanțe diferite în citoplasmă, cum ar fi, de exemplu, la amfibieni, transcripția nu este activată imediat. Sinteza ARN în ele începe în stadiul blastulei timpurii. Dimpotrivă, la mamifere, sinteza ARN începe deja în stadiul a două blastomere.
În perioada de clivaj se formează ARN și proteine, similare celor sintetizate în timpul oogenezei. Acestea sunt în principal histone, proteine ale membranei celulare și enzime necesare diviziunii celulare. Aceste proteine sunt folosite imediat împreună cu proteinele stocate mai devreme în citoplasma ovocitelor. Odată cu aceasta, în perioada de zdrobire, este posibilă sinteza proteinelor, ceea ce nu era acolo înainte. Acest lucru este susținut de date privind prezența diferențelor regionale în sinteza ARN și a proteinelor între blastomeri. Uneori, aceste ARN și proteine intră în acțiune într-o etapă ulterioară.
Un rol important în zdrobire îl joacă diviziunea citoplasmei - citotomie. Are o semnificație morfogenetică deosebită, deoarece determină tipul de zdrobire. În procesul de citotomie, se formează mai întâi o constricție cu ajutorul unui inel contractil de microfilamente. Asamblarea acestui inel are loc sub influența directă a polilor fusului mitotic. După citotomie, blastomerele ouălor oligolecite rămân conectate între ele doar prin punți subțiri. În acest moment este cel mai ușor să le separați. Acest lucru se datorează faptului că citotomia duce la o reducere a zonei de contact dintre celule din cauza suprafeței limitate a membranelor.
Imediat după citotomie, începe sinteza de noi secțiuni ale suprafeței celulare, zona de contact crește, iar blastomerele încep să se atingă strâns. Brazdele de clivaj se desfășoară de-a lungul limitelor dintre secțiunile individuale ale ovoplasmei, reflectând fenomenul de segregare ovoplasmatică. Din acest motiv, citoplasma diferitelor blastomere diferă în compoziția chimică.
Zdrobire - concept și tipuri. Clasificarea și caracteristicile categoriei „Zdrobire” 2017, 2018.
La sfârșitul secolului XII - începutul secolului XIII. pe baza renașterii sociale și economice generale a Germaniei, s-au făcut schimbări importante în structura politică a imperiului: fostele regiuni feudale (ducate, arhiepiscopii) s-au transformat în state aproape complet independente.... .
FERTILIZARE Cursul 8 Fertilizarea este stimularea unui ovul de către un spermatozoid să se dezvolte, transferând simultan materialul ereditar al tatălui în ovul. În procesul de fertilizare, spermatozoidul fuzionează cu ovulul, în timp ce nucleul haploid ....
PROCESE PREGĂTITORII PRELEGERE Nr.4 Spălarea mineralelor Spălarea este utilizată în îmbogățirea zăcămintelor de placer de metale rare și prețioase, minereuri de metale feroase, fosforiți, caolini, materiale de construcție (nisip, piatră spartă),...
După convergența pronucleilor feminin și masculin, care durează aproximativ 12 ore la mamifere, o zigot- embrion unicelular. Deja în stadiul de zigot, zonele prezumtive (lat. presumptio - probabilitate, presupunere) sunt dezvăluite ca surse de dezvoltare a secțiunilor corespunzătoare ale blastulei, din care se formează ulterior straturile germinale.
Orez. Zigotul uman în stadiul de convergență a nucleilor masculin și feminin (pronuclei): (după B.P. Hvatov).
1 - miez feminin; 2 - nucleul masculin.
Clivajul și formarea blastulei
Clivaj - diviziunea mitotică secvențială a zigotului în celule (blastomere) fără creșterea celulelor fiice la dimensiunea mamei.
Blastomerele rezultate rămân unite într-un singur organism al embrionului. La zigot se formează un fus mitotic între centrioli care se îndepărtează spre poli, introdus de spermatozoid. Pronucleii intră în stadiul de profază cu formarea unui set diploid combinat (metoda de detectare a zonelor prezumtive a fost propusă de embriologul german Vogt) de cromozomi de ovul și spermatozoizi. După ce a trecut prin toate celelalte faze ale diviziunii mitotice, zigotul este împărțit în două celule fiice - blastomere. Datorită absenței virtuale a perioadei G 1, în care celulele formate ca urmare a diviziunii cresc, celulele sunt mult mai mici decât celula mamă, prin urmare, dimensiunea embrionului în ansamblu în această perioadă, indiferent de numărul celulelor sale constitutive, nu depășește dimensiunea celulei originale - zigotul. Toate acestea au făcut posibilă denumirea procesului descris de zdrobire (adică măcinare) și celulele formate în procesul de zdrobire - blastomeri.
Scindarea zigotului uman începe la sfârșitul primei zile și este caracterizată ca fiind complet neuniformă asincronă. În primele zile, se întâmplă încet. Prima scindare (diviziune) a zigotului este finalizată după 30 de ore, rezultând formarea a 2 blastomeri acoperiți cu o membrană de fertilizare. Stadiul a două blastomere este urmat de stadiul a trei blastomere.
De la prima zdrobire a zigotului, se formează două tipuri de blastomeri - „întunecat” și „luminos”. „Ușoare”, mai mici, blastomerele sunt zdrobite mai repede și sunt aranjate într-un singur strat în jurul „întunericului” mare, care se află în mijlocul embrionului. Din blastomerele „ușoare” de suprafață, ia naștere ulterior un trofoblast, care conectează embrionul cu corpul mamei și asigură nutriția acestuia. Blastomerele interne, „întunecate”, formează embrioblastul, din care se formează corpul embrionului și unele organe extraembrionare (amnion, sacul vitelin, alantoide).
Începând de la trei zile, fragmentarea se desfășoară mai rapid, iar în a 4-a zi embrionul este format din 7-12 blastomeri. După 50-60 de ore, se formează o acumulare densă de celule - morula, iar în a 3-4-a zi începe formarea unui blastocist - o veziculă goală umplută cu lichid (Fig.

Orez. Embrionul uman în stadiile incipiente de dezvoltare (după Hertig și Rock).
A - stadiul a două blastomere; B - blastocist; I - embrioblast, 2 - trofoblast; 3 - cavitatea blastochistului.
Blastocistul se deplasează prin oviduct către uter în decurs de 3 zile și după 4 zile. intră în uter. Blastocistul este liber în cavitatea uterină timp de 2 zile (zilele 5 și 6), iar această etapă este denumită blastocist liber. Până în acest moment, blastocistul crește din cauza creșterii numărului de blastomere - celule embrioblastice și trofoblastice - până la 100 sau mai mult datorită absorbției crescute a secreției glandelor uterine de către trofoblast și, de asemenea, datorită producției active de fluid de către trofoblastul însuși (Fig.
Embrioblastul este situat sub forma unui mănunchi de celule germinale („mănunchi germinativ”), care este atașat din interior de trofoblast la unul dintre polii blastocistului și începe implantarea.
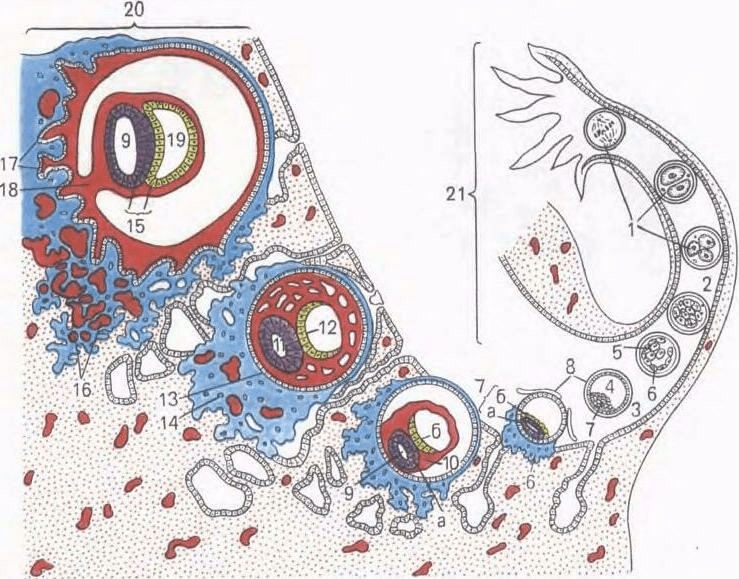
Orez. 37. Clivajul, gastrulația și implantarea embrionului uman (schemă).
1 - zdrobire, 2 - morula; 3 - blastocist; 4 - cavitatea blastocistului; 5 - embrioblast; 6 - trofoblast; 7 - nodul germinativ: a - epiblast, b - hipoblast: 8 - membrana de fertilizare; 9 - veziculă amniotică (ectodermică); 10 - mezoderm extraembrionar; II - ectoderm; 12 - endoderm; 13 - citotrofoblast; 14 - simplastotrofoblast; 15 - disc germinativ; 16 - goluri cu sânge matern; 17 - corion; 18 - picior amniotic; 19 - veziculă de gălbenuș; 20 - membrana mucoasă a uterului; 21 - oviduct.
Fertilizare este procesul de fuziune a celulelor sexuale. Celula diploidă formată în urma fertilizării - zigotul - este etapa inițială în dezvoltarea unui nou organism. Procesul de fertilizare constă din trei faze succesive:
a) convergenţa gameţilor(Hamonii (hormonii gameților), pe de o parte, activează mișcarea spermatozoizilor, iar pe de altă parte, lipirea acestora.) În momentul contactului spermatozoidului cu membrana celulei ovulului, are loc o reacție acrozomală, în timpul căreia , sub acțiunea enzimelor proteolitice ale acrozomului, membranele de ou se dizolvă. Mai mult, membranele plasmatice ale ovulului și spermatozoizilor fuzionează și prin puntea citoplasmatică rezultată a citoplasmei ambilor gameți sunt combinate. Apoi nucleul și centriolul spermatozoidului trec în citoplasma oului, iar membrana spermatozoidului este încorporată în membrana celulei ou. Partea coadă a spermatozoizilor la majoritatea animalelor intră și ea în ovul, dar apoi se separă și se dizolvă, fără a juca niciun rol în dezvoltarea ulterioară;
b) activarea ouluiDatorită faptului că secțiunea membranei spermatozoidului este permeabilă la ionii de sodiu, aceștia din urmă încep să intre în ovul, modificând potențialul de membrană al celulei. Apoi, sub forma unei unde care se propagă din punctul de contact al gameților, are loc o creștere a conținutului de ioni de calciu, urmată de dizolvarea granulelor corticale sub formă de undă. Enzimele specifice eliberate în același timp contribuie la desprinderea membranei gălbenușului; se intareste, este coaja fertilizarii. Toate procesele descrise sunt așa-numita reacție corticală.;
c) fuziunea gameților sau singamieOvulul în momentul întâlnirii cu spermatozoizii se află de obicei într-unul dintre etapele meiozei, blocat de un factor specific. La majoritatea vertebratelor, acest bloc apare în stadiul metafazei II; la multe nevertebrate, precum și la trei specii de mamifere (cai, câini și vulpi), blocul apare în stadiul de diakineză. În cele mai multe cazuri, blocul meiotic este îndepărtat după activarea oului din cauza fertilizării. În timp ce meioza este finalizată în ovul, nucleul spermatozoizilor care a pătruns în el este modificat. Ea ia forma unei interfaze și apoi a unui nucleu de profază. În acest timp, ADN-ul se dublează și pronucleul masculin primește cantitatea de material ereditar corespunzătoare p2c, adică. conține un set haploid de cromozomi reduplicați. Nucleul celulei ou, care a terminat meioza, se transformă într-un pronucleu feminin, dobândind și n2c. Ambii pronuclei fac mișcări complexe, apoi se apropie și se contopesc (sincarion), formând o placă metafază comună. Acesta, de fapt, este momentul fuziunii finale a gameților - singamia. Prima diviziune mitotică a zigotului are ca rezultat formarea a două celule embrionare (blastomere) cu un set de cromozomi 2n2c în fiecare.
zigot - o celulă diploidă (conținând un set complet dublu de cromozomi) rezultată în urma fecundației (fuziunea unui ovul și spermatozoizii). Zigotul este o celulă totipotentă (adică capabilă să producă orice altă celulă).
La om, prima diviziune mitotică a zigotului are loc la aproximativ 30 de ore după fertilizare, ceea ce se datorează proceselor complexe de pregătire pentru primul act de zdrobire. Celulele formate ca urmare a zdrobirii zigotului se numesc
blastomeri. Primele diviziuni ale unui zigot se numesc „zdrobire” deoarece celula este zdrobită: după fiecare diviziune, celulele fiice devin din ce în ce mai mici și nu există nicio etapă de creștere a celulelor între diviziuni.
Despărțirea - aceasta este o serie de diviziuni mitotice succesive ale zigotului și ale altor blastomere, care se termină cu formarea unui embrion multicelular - blastula.Între diviziunile succesive, creșterea celulară nu are loc, dar ADN-ul este neapărat sintetizat. Toți precursorii ADN și enzimele necesare se acumulează în timpul oogenezei. În primul rând, blastomerele sunt adiacente unul altuia, formând un grup de celule numite morula. Apoi se formează o cavitate între celule - blastocoel, umplut cu lichid. Celulele sunt împinse la periferie, formând peretele blastulei - blastoderm. Dimensiunea totală a embrionului până la sfârșitul clivajului în stadiul de blastula nu depășește dimensiunea zigotului. Principalul rezultat al perioadei de zdrobire este transformarea zigotului în embrion multicelular unilamelar.
Morfologia strivirii.De regulă, blastomerele sunt aranjate într-o ordine strictă unul față de celălalt și axa polară a oului. Ordinea sau metoda de zdrobire depinde de cantitatea, densitatea și distribuția gălbenușului în ou. Conform regulilor lui Sachs-Hertwig, nucleul celular tinde să fie situat în centrul citoplasmei fără gălbenuș, iar axul diviziunii celulare - în direcția cea mai mare a acestei zone.
În oligo- și mesolecital La ouă, clivajul este complet sau holoblastic. Acest tip de strivire se găsește la lamprede, unii pești, toți amfibieni, precum și la marsupiale și mamiferele placentare. La zdrobire completă, planul primei diviziuni corespunde planului de simetrie bilaterală. Planul celei de-a doua diviziuni merge perpendicular pe planul primei. Ambele brazde ale primelor două diviziuni sunt meridiane, adică. începe de la polul animal și se extinde la polul vegetativ. Celula ou este împărțită în patru blastomere mai mult sau mai puțin egale ca dimensiuni. Planul celei de-a treia diviziuni se desfășoară perpendicular pe primele două în direcția latitudinală. După aceea, în ouăle mesolecitale în stadiul de opt blastomere se manifestă zdrobirea neuniformă. Pe polul animal sunt patru blastomeri mai mici - micromere, pe polul vegetativ - patru mai mari - macromere. Apoi diviziunea merge din nou în planurile meridiane și apoi din nou în latitudinal.
În polilecitalîn ouăle peștilor osoși, reptilelor, păsărilor, precum și mamiferelor monotreme, fragmentarea este parțială sau meroblastică, adică acoperă doar citoplasma lipsită de gălbenuș. Este situat sub forma unui disc subțire la polul animalului, de aceea acest tip de zdrobire se numește discoidal. La caracterizarea tipului de zdrobire se ia în considerare și poziția relativă și rata de divizare a blastomerilor. Dacă blastomerele sunt dispuse în rânduri unul deasupra celuilalt de-a lungul razelor, zdrobirea se numește radială. Este tipic pentru cordate și echinoderme. În natură, există și alte variante de aranjare spațială a blastomerelor în timpul zdrobirii, ceea ce determină astfel de tipuri de ele ca spirală la moluște, bilaterală la ascaris, anarhică la meduze.
S-a remarcat o relație între distribuția gălbenușului și gradul de sincronism în diviziunea blastomerelor animale și vegetative. În ouăle oligolecite ale echinodermelor, clivajul este aproape sincron; în celulele ouă mezocitetale, sincronia este perturbată după a treia diviziune, deoarece blastomerele vegetative se divid mai lent din cauza cantității mari de gălbenuș. În formele cu clivaj parțial, diviziunile sunt asincrone de la bun început, iar blastomerele care ocupă o poziție centrală se divid mai repede.
La sfârșitul zdrobirii, se formează o blastula. Tipul de blastula depinde de tipul de zdrobire și, prin urmare, de tipul de ou.
Caracteristicile proceselor molecular-genetice și biochimice în timpul zdrobirii.După cum sa menționat mai sus, ciclurile mitotice în timpul perioadei de clivaj sunt mult scurtate, mai ales la început.
De exemplu, întregul ciclu de fisiune în ouăle de arici de mare durează 30-40 de minute, în timp ce durata fazei S este de doar 15 minute. Perioadele GI și G2 sunt practic absente, deoarece aportul necesar de toate substanțele a fost creat în citoplasma celulei ou și cu cât este mai mare, cu atât este mai mare. Înainte de fiecare diviziune are loc sinteza ADN-ului și a histonelor.
Rata cu care furculița de replicare se mișcă de-a lungul ADN-ului în timpul clivajului este normală. În același timp, există mai multe puncte de inițiere în ADN-ul blastomerilor decât în celulele somatice. Sinteza ADN-ului are loc în toți repliconii simultan, sincron. Prin urmare, timpul de replicare a ADN-ului în nucleu coincide cu timpul de dublare a unuia, în plus, scurtat, replicon. S-a demonstrat că atunci când nucleul este îndepărtat din zigot, are loc clivajul, iar embrionul în dezvoltare ajunge aproape în stadiul de blastulă. Dezvoltarea ulterioară se oprește.
La începutul clivajului, alte tipuri de activitate nucleară, cum ar fi transcripția, sunt practic absente. În diferite tipuri de ouă, transcripția genelor și sinteza ARN încep în diferite etape. În cazurile în care există multe substanțe diferite în citoplasmă, cum ar fi, de exemplu, la amfibieni, transcripția nu este activată imediat. Sinteza ARN în ele începe în stadiul blastulei timpurii. Dimpotrivă, la mamifere, sinteza ARN începe deja în stadiul a două blastomere.
În perioada de clivaj se formează ARN și proteine, similare celor sintetizate în timpul oogenezei. Acestea sunt în principal histone, proteine ale membranei celulare și enzime necesare diviziunii celulare. Aceste proteine sunt folosite imediat împreună cu proteinele stocate mai devreme în citoplasma ovocitelor. Odată cu aceasta, în perioada de zdrobire, este posibilă sinteza proteinelor, ceea ce nu era acolo înainte. Acest lucru este susținut de date privind prezența diferențelor regionale în sinteza ARN și a proteinelor între blastomeri. Uneori, aceste ARN și proteine intră în acțiune într-o etapă ulterioară.
Un rol important în zdrobire îl joacă diviziunea citoplasmei - citotomie. Are o semnificație morfogenetică deosebită, deoarece determină tipul de zdrobire. În procesul de citotomie, se formează mai întâi o constricție cu ajutorul unui inel contractil de microfilamente. Asamblarea acestui inel are loc sub influența directă a polilor fusului mitotic. După citotomie, blastomerele ouălor oligolecite rămân conectate între ele doar prin punți subțiri. În acest moment sunt cel mai ușor de separat. Acest lucru se datorează faptului că citotomia duce la o scădere a zonei de contact dintre celule din cauza suprafeței limitate a membranelor. Imediat după citotomie, începe sinteza de noi secțiuni ale suprafeței celulare, zona de contact crește și blastomerele încep să apară. atinge strâns. Brazdele de clivaj se desfășoară de-a lungul limitelor dintre secțiunile individuale ale ovoplasmei, reflectând fenomenul de segregare ovoplasmatică. Prin urmare, citoplasma diferitelor blastomere diferă în compoziția chimică.
Caracteristicile și semnificația principalelor etape ale dezvoltării embrionare: gastrulația, histo- și organogeneza. Formarea de embrioni cu 2 și 3 straturi. Metode de formare a mezodermului. Derivați ai straturilor germinale. Mecanisme de reglare a acestor procese la nivel de gene și celulare.
Histogenie- (din greacă. histos - tissue it ... geneza), un ansamblu de procese care s-a dezvoltat în filogeneză, asigurând formarea, existența și refacerea țesuturilor cu caracteristicile lor inerente specifice organelor în ontogeneza organismelor pluricelulare. Caracteristici. În organism, țesuturile se dezvoltă din anumite rudimente embrionare (straturi germinale derivate) formate ca urmare a proliferării, mișcării (mișcări morfogenetice) și aderării celulelor embrionare în stadiile incipiente ale dezvoltării sale în procesul de organogeneză. Ființe, factorul lui G. - diferențierea celulelor determinate ducând la apariția diferitelor morfol. și physiol. tipuri de celule care sunt distribuite în mod regulat în organism. Uneori G. este urmată de formarea substanţei intercelulare. Un rol important în determinarea direcției lui G. revine interacțiunilor de contact intercelular și influențelor hormonale. Ansamblul de celule care efectuează un anumit G., este subîmpărțit într-un număr de grupuri: celule ancestrale (stem) capabile de diferențiere și completare a pierderii propriului fel prin diviziune; celulele progenitoare (așa-numitele celule semi-stem) - se diferențiază, dar își păstrează capacitatea de a se diviza; difer matur. celule. G. reparatorie în perioada postnatală stă la baza refacerii țesuturilor deteriorate sau parțial pierdute. Calități, modificările lui G. pot duce la apariția și creșterea unei tumori.
Organogeneza(din grecescul organon - organ, geneza - dezvoltare, educație) - procesul de dezvoltare, sau formare, a organelor în embrionul uman și al animalelor. Organogeneza urmează perioadele anterioare ale dezvoltării embrionare (vezi Embrion) - zdrobirea ouălor, gastrularea și are loc după ce principalele rudimente (marcaje) ale organelor și țesuturilor sunt izolate. Organogeneza se desfășoară în paralel cu histogeneza (vezi) sau dezvoltarea țesuturilor. Spre deosebire de țesuturi, fiecare dintre ele având ca sursă unul dintre rudimentele embrionare, organele, de regulă, apar cu participarea mai multor (de la două la patru) rudimente diferite (vezi Straturile germinale), dând naștere la diferite componente tisulare ale organ. De exemplu, ca parte a peretelui intestinal, epiteliul care căptușește cavitatea organului și glandele se dezvoltă din stratul germinal interior - endodermul (vezi), țesutul conjunctiv cu vase și țesut muscular neted - din mezenchim (vezi), mezoteliul care acoperă membrana seroasă a intestinului, - din frunza viscerală a splanhnotomului, adică frunza germinativă mijlocie - mezodermul, iar nervii și ganglionii organului - din rudimentul neural. Pielea este formată cu participarea stratului germinal exterior - ectoderm (vezi), din care se dezvoltă epiderma și derivații săi (păr, glande sebacee și sudoripare, unghii etc.) și dermatoame, din care ia naștere mezenchimul, diferențiându-se în baza țesutului conjunctiv al pielii (derma). Nervii și terminațiile nervoase din piele, ca și în alte părți, sunt derivați ai germenului neural. Unele organe sunt formate dintr-un singur germ, de exemplu, os, vase de sânge, ganglioni limfatici - din mezenchim; cu toate acestea, și aici, derivate ale rudimentului sistemului nervos - fibrele nervoase - cresc în anlage și se formează terminații nervoase.
Dacă histogeneza constă în principal în reproducerea și specializarea celulelor, precum și în formarea de substanțe intercelulare și alte structuri necelulare, atunci principalele procese care stau la baza organogenezei sunt formarea straturilor germinale de pliuri, proeminențe, proeminențe, îngroșări, neuniforme. creșterea, fuziunea sau divizarea (separarea), precum și germinarea reciprocă a diferitelor semne de carte. La om, organogeneza începe la sfârșitul săptămânii a 3-a și se termină în termeni generali în luna a 4-a de dezvoltare intrauterină. Cu toate acestea, dezvoltarea unui număr de organe provizorii (temporare) ale embrionului - corion, amnios, sacul vitelin - începe deja la sfârșitul primei săptămâni, iar unele organe definitive (finale) se formează mai târziu decât altele (de exemplu, limfa). ganglionii – începând din ultimele luni de dezvoltare intrauterină și până la debutul pubertății).
Gastrulatie -un embrion cu un singur strat - blastula - se transformă într-un multistrat - cu două sau trei straturi, numit gastrula (din grecescul gaster - stomac în sens diminutiv).
În cordate primitive, de exemplu, în lancetă, un blastoderm omogen cu un singur strat în timpul gastrulației este transformat într-un strat germinal exterior, ectodermul, și într-un strat germinal interior, endodermul. Endodermul formează intestinul primar cu o cavitate în interior, gastrocelul. Deschiderea care duce la gastrocel se numește blastopor sau gura primară. Două straturi germinale sunt caracteristicile morfologice definitorii ale gastrulației. Existența lor la un anumit stadiu de dezvoltare la toate animalele pluricelulare, de la celenterate până la vertebratele superioare, ne permite să ne gândim la omologia straturilor germinale și la unitatea originii tuturor acestor animale. La vertebrate, pe lângă cele două menționate, în timpul gastrulației, se formează un al treilea strat germinal - mezodermul, care ocupă un loc între ecto- și endoderm. Dezvoltarea stratului germinal mijlociu, care este un cordomezoderm, este o complicație evolutivă a fazei de gastrulație la vertebrate și este asociată cu o accelerare a dezvoltării lor în stadiile incipiente ale embriogenezei. La cordatele mai primitive, cum ar fi lanceta, cordomezodermul se formează de obicei la începutul fazei următoare după gastrulație - organogeneză. Schimbarea timpului de dezvoltare a unor organe față de altele la descendenți în comparație cu grupurile ancestrale este o manifestare a heterocroniei. Schimbările în momentul formării celor mai importante organe în cursul evoluției nu sunt neobișnuite.
Procesul de gastrulare se caracterizează prin transformări celulare importante, cum ar fi mișcările direcționate ale grupurilor și ale celulelor individuale, reproducerea selectivă și sortarea celulelor, începutul citodiferențierii și interacțiunile de inducție.
Metode de gastrularediferit. Se disting patru tipuri de mișcări celulare direcționate spațial, ducând la transformarea embrionului dintr-un singur strat într-unul multistrat.
Invaginatie- invaginarea uneia dintre secțiunile blastodermului spre interior ca un strat întreg. În lancetă, celulele polului vegetativ se invaginează; la amfibieni, invaginarea apare la granița dintre polii animal și vegetativ în regiunea semilunei cenușii. Procesul de invaginare este posibil numai în ouă cu o cantitate mică sau medie de gălbenuș.
epibolie- murdărirea cu celule mici ale polului animal de celule mai mari, întârziate în rata de diviziune și mai puțin mobile ale polului vegetativ. Acest proces este exprimat clar la amfibieni.
Denumirea- stratificarea celulelor blastodermice în două straturi situate unul deasupra celuilalt. Delaminarea poate fi observată în discoblastula embrionilor cu un tip parțial de strivire, cum ar fi reptilele, păsările și mamiferele ovipare. Delaminarea se manifestă în embrioblastul mamiferelor placentare, ducând la formarea de hipoblast și epiblast.
Imigrare- mișcarea unor grupuri sau celule individuale care nu sunt unite într-un singur strat. Imigrația are loc la toți embrionii, dar este cea mai caracteristică fazei a doua a gastrulației la vertebratele superioare. În fiecare caz specific de embriogeneză, de regulă, sunt combinate mai multe metode de gastrulare.
Morfologia gastrulației.În regiunea blastulei, din materialul celular al căruia, în timpul gastrulației și organogenezei timpurii (neurulare), se formează de obicei straturi germinale și organe complet definite. Invaginarea începe la polul vegetativ. Datorită diviziunii mai rapide, celulele polului animal cresc și împing celulele polului vegetativ în blastula. Acest lucru este facilitat de o schimbare a stării citoplasmei în celulele care formează buzele blastoporului și adiacente acestora. Datorită invaginării, blastocelul scade și gastrocelul crește. Concomitent cu dispariția blastocelului, ectodermul și endodermul intră în contact strâns. În lancetă, la fel ca în toate deuterostomele (include tipul echinoderm, tipul cordat și alte câteva tipuri mici de animale), regiunea blastoporului se transformă în partea coadă a organismului, spre deosebire de protostomi, în care blastoporul corespunde. la partea capului. Deschiderea gurii în deuterostomi se formează la capătul embrionului opus blastoporului. Gastrularea la amfibieni are multe în comun cu gastrulația lanceletei, dar deoarece gălbenușul din ouăle lor este mult mai mare și este situat în principal la polul vegetativ, blastomerii mari de amfiblastula nu sunt capabili să se bombanească spre interior.Invaginatie merge putin diferit. La granița dintre polii animal și vegetativ din regiunea secerului gri, celulele sunt mai întâi puternic atrase spre interior, luând forma „în formă de balon”, apoi trag celulele stratului de suprafață al blastulei împreună cu lor. Apar un șanț semilună și o buză blastopor dorsală. În același timp, celulele mai mici ale polului animal, divându-se mai repede, încep să se deplaseze spre polul vegetativ. În regiunea buzei dorsale, ele se ridică și se invaginează, iar celulele mai mari cresc pe părțile laterale și pe partea opusă șanțului în formă de seceră. Apoi procesulepibolie duce la formarea buzelor laterale și ventrale ale blastoporului. Blastoporul se închide într-un inel, în interiorul căruia celulele luminoase mari ale polului vegetativ sunt vizibile de ceva timp sub forma așa-numitului dop de gălbenuș. Mai târziu, ele sunt complet scufundate în interior, iar blastoporul se îngustează. Folosind metoda de marcare cu coloranți vitali (vitali) la amfibieni au fost studiate în detaliu mișcările celulelor blastulelor în timpul gastrulației, apoi în interiorul organelor în sine. Se știe că la anure materialul notocordului și mezodermului prezumtiv la stadiul de blastula nu se află pe suprafața sa, ci în straturile interioare ale peretelui amfiblastulei, totuși, aproximativ la nivelurile prezentate în figură. O analiză a etapelor incipiente ale dezvoltării amfibienilor ne permite să concluzionăm că segregarea ovoplasmatică, care se manifestă în mod clar în ou și zigot, are o mare importanță în determinarea soartei celulelor care au moștenit o anumită zonă a citoplasmei. Gastrularea la embrionii cu un tip de clivaj și dezvoltare meroblastic are propriile sale caracteristici. La păsări, începe după zdrobire și formarea blastulei în timpul trecerii embrionului prin oviduct. În momentul în care oul este depus, embrionul este deja format din mai multe straturi: stratul superior se numește epiblast, stratul inferior se numește hipoblast primar. Între ele este un decalaj îngust - blastocelul. Apoi se formează un hipoblast secundar, a cărui metodă de formare nu este complet clară. Există dovezi că celulele germinale primare își au originea în hipoblastul primar al păsărilor, în timp ce cel secundar formează endodermul extraembrionar. Formarea hipoblastului primar și secundar este considerată ca un fenomen premergător gastrulației. Principalele evenimente ale gastrulației și formarea finală a celor trei straturi germinale încep după ovipunerea cu debutul incubației. Există o acumulare de celule în partea posterioară a epiblastului ca urmare a vitezei neuniforme de diviziune celulară și a mișcării lor din părțile laterale ale epiblastului spre centru, unul către celălalt. Se formează o așa-numită dungă primară, care se extinde spre capătul capului. În centrul benzii primare, se formează o canelură primară, iar de-a lungul marginilor, creste primare. La capătul capului benzii primare, apare o îngroșare - nodul Hensen, iar în el - fosa primară. Când celulele epiblastice intră în șanțul primar, forma lor se schimbă. Ele seamănă cu forma celulelor „în formă de balon” ale gastrulei amfibienilor. Aceste celule devin apoi stelate și se scufundă sub epiblast pentru a forma mezodermul. Endodermul se formează pe baza hipoblastului primar și secundar cu adăugarea unei noi generații de celule endodermice migrând din straturile superioare, blastodermul. Prezența mai multor generații de celule endodermice indică prelungirea perioadei de gastrulație în timp. O parte din celulele care migrează din epiblast prin nodul Hensen formează viitoarea notocordă. Concomitent cu inițierea și alungirea coardei, nodul Hensen și stria primară dispar treptat în direcția de la capătul anterior spre cel caudal. Aceasta corespunde îngustării și închiderii blastoporului. Pe măsură ce dâra primară se contractă, ea lasă în urmă secțiunile formate ale organelor axiale ale embrionului în direcția de la cap la secțiunile cozii. Pare rezonabil să se considere mișcările celulelor din embrionul de pui ca epibol omolog, iar stria primară și nodul Hensen ca omoloage cu blastoporul din buza dorsală a gastrulei amfibiene. Este interesant de observat că celulele embrionilor de mamifere, în ciuda faptului că la aceste animale ouăle au o cantitate mică de gălbenuș, iar fragmentarea este completă, în faza de gastrulare păstrează mișcările caracteristice embrionilor de reptile și păsări. Acest lucru confirmă ideea originii mamiferelor dintr-un grup ancestral ale cărui ouă erau bogate în gălbenuș.
Caracteristicile etapei de gastrulație.Gastrulația este caracterizată printr-o varietate de procese celulare. Reproducerea mitotică a celulelor continuă și are o intensitate diferită în diferite părți ale embrionului. Cu toate acestea, cea mai caracteristică trăsătură a gastrulației este mișcarea maselor celulare. Aceasta duce la o schimbare a structurii embrionului și la transformarea acestuia din blastula în gastrula. Celulele sunt sortate în funcție de apartenența lor la diferite straturi germinale, în interiorul cărora se „recunoaște” reciproc. Faza de gastrulare marchează începutul citodiferențierii, ceea ce înseamnă o tranziție la utilizarea activă a informațiilor biologice ale propriului genom. Unul dintre regulatorii activității genetice este compoziția chimică diferită a citoplasmei celulelor embrionare, care se stabilește ca urmare a segregării ovoplasmatice. Deci, celulele ectodermice ale amfibienilor au o culoare închisă datorită pigmentului care a intrat în ele de la polul animal al oului, iar celulele endodermice sunt ușoare, deoarece provin de la polul vegetativ al oului. În timpul gastrulației, rolul inducției embrionare este foarte mare. S-a demonstrat că apariția striei primare la păsări este rezultatul unei interacțiuni inductive între hipoblast și epiblast. Hipoblastul are polaritate. O modificare a poziției hipoblastului față de epiblast provoacă o schimbare a orientării striei primitive. Toate aceste procese sunt descrise în detaliu în acest capitol. Trebuie remarcat faptul că astfel de manifestări ale integrității embrionului, cum ar fi determinarea, reglarea embrionară și integrarea, sunt inerente în timpul gastrulației în aceeași măsură ca și în timpul clivajului.
Formarea mezodermului - La toate animalele, cu excepția cavităților intestinale, în legătură cu gastrulația (în paralel cu aceasta sau în etapa următoare, datorită gastrulației), apare și un al treilea strat germinativ, mezodermul. Aceasta este o colecție de elemente celulare care se află între ectoderm și endoderm, adică în blastocel. Asa. Astfel, embrionul devine nu în două straturi, ci în trei straturi. La vertebratele superioare, structura cu trei straturi a embrionilor apare deja în procesul de gastrulare, în timp ce la cordatele inferioare și la toate celelalte tipuri, ca urmare a gastrulației în sine, se formează un embrion cu două straturi.
Pot fi stabilite două moduri fundamental diferite de apariție a mezodermului:teloblastic, caracteristic Protostomiei, și enterocelic, caracteristic Deute-rosiomiei. în protostome în timpul gastrulației, la granița dintre ectoderm și endoderm, pe părțile laterale ale blastoporului, există deja două celule mari care separă celulele mici de ele însele (din cauza diviziunilor). Astfel, se formează stratul mijlociu - mezodermul. Teloblastele, dând noi și noi generații de celule mezodermice, rămân la capătul posterior al embrionului. Din acest motiv, această metodă de formare a mezodermului se numește teloblastică (din grecescul telos - sfârșit).
Cu metoda enterocelului, totalitatea celulelor mezodermului emergent apare sub formă de proeminențe asemănătoare buzunarului intestinului primar (protruzie a pereților acestuia în blastocel). Aceste proeminențe, în interiorul cărora intră părți din cavitatea intestinală primară, sunt izolate de intestin și separate de acesta sub formă de saci. Cavitatea sacilor se transformă într-un întreg, adică într-o cavitate corporală secundară, sacii celomici pot fi subdivizați în segmente ale stratului germinal mijlociu nu reflectă întreaga varietate de variații și abateri care sunt strict naturale pentru grupuri individuale de animale. . Similar cu teloblastul, dar numai în exterior, metoda de formare a mezodermului nu este prin divizarea teloblastelor, ci prin apariția unui primordiu dens nepereche (grup de celule) la marginile blastoporului, care ulterior se împarte în două dungi simetrice de celule. Prin metoda enterocelului, primordiul mezodermului poate fi împerecheat sau neîmperecheat; în unele cazuri, se formează doi saci celomici simetrici, în timp ce în altele, se formează mai întâi un sac celomic comun, care ulterior se împarte în două jumătăți simetrice.
Derivați ai straturilor germinale.Soarta ulterioară a celor trei straturi germinale este diferită.
Din ectoderm se dezvoltă: tot țesutul nervos; straturile exterioare ale pielii și derivatele acesteia (păr, unghii, smalț dentar) și parțial membrana mucoasă a cavității bucale, cavitățile nazale și anus.
Endodermul dă naștere căptușelii întregului tub digestiv - de la cavitatea bucală până la anus - și tuturor derivaților săi, adică. timus, tiroida, glande paratiroide, trahee, plamani, ficat si pancreas.
Din mezoderm se formează: toate tipurile de țesut conjunctiv, țesut osos și cartilaj, sânge și sistemul vascular; toate tipurile de țesut muscular; sistemele excretor și reproducător, stratul dermic al pielii.
La un animal adult, există foarte puține organe de origine endodermică care nu conțin celule nervoase derivate din ectoderm. Fiecare organ important conține, de asemenea, derivați ai mezodermului - vase de sânge, sânge și adesea mușchi, astfel încât izolarea structurală a straturilor germinale să fie păstrată doar în stadiul formării lor. Deja la începutul dezvoltării lor, toate organele capătă o structură complexă și includ derivați ai tuturor straturilor germinale.
Începutul unui nou organism este dat de un ovul fecundat (cu excepția cazurilor de partenogeneză și reproducere vegetativă). Fertilizarea este procesul de fuziune a două celule germinale (gameți) unul cu celălalt, în timpul căruia sunt îndeplinite două funcții diferite: sexuală (combinând genele a doi indivizi parentali) și reproductivă (apariția unui nou organism). Prima dintre aceste funcții include transferul de gene de la părinți la urmași, a doua - inițierea în citoplasma oului a acelor reacții și mișcări care permit dezvoltarea continuă. Ca rezultat al fertilizării, un set dublu (2p) de cromozomi este restaurat în ovul. Centrozomul, introdus de spermatozoizi, după dublare formează un fus de fisiune, iar zigotul intră în stadiul I de embriogeneză – stadiul strivirii. Ca urmare a mitozei, din zigot se formează 2 celule fiice - blastomere.
Perioada prezigotică
Perioada de dezvoltare prezigotică este asociată cu formarea gameților (gametogeneza). Formarea ovocitelor începe la femei chiar înainte de a se naște și se finalizează pentru fiecare ovocit dat numai după fecundarea acestuia. Până la naștere, fătul feminin din ovare conține aproximativ două milioane de ovocite de ordinul întâi (acestea sunt încă celule diploide), iar doar 350 - 450 dintre ele vor ajunge în stadiul de ovocite de ordinul doi (celule haploide), transformându-se. în ouă (unul câte unul în timpul unui ciclu menstrual). ). Spre deosebire de femei, celulele germinale din testicule (testiculele) la bărbați încep să se formeze abia odată cu debutul pubertății. Durata perioadei de formare a spermatozoizilor este de aproximativ 70 de zile; pentru un gram de greutate a testiculelor, numărul de spermatozoizi este de aproximativ 100 de milioane pe zi.
Fertilizare
Fertilizarea - fuziunea unei celule reproductive masculine (sperma) cu o femelă (ou, ovul), ducând la formarea unui zigot - un nou organism unicelular. Semnificația biologică a fertilizării este unificarea materialului nuclear al gameților masculini și feminini, ceea ce duce la unificarea genelor paterne și materne, refacerea setului diploid de cromozomi, precum și activarea oului, adică stimularea acestuia pentru dezvoltarea embrionară. Conexiunea ovulului cu spermatozoidul are loc de obicei în partea în formă de pâlnie a trompei uterine în primele 12 ore după ovulație.
Lichidul seminal, care intră în vaginul femeii în timpul actului sexual, conține de obicei de la 60 la 150 de milioane de spermatozoizi, care, datorită mișcărilor cu o viteză de 2-3 mm pe minut, contracțiilor ondulatorii constante ale uterului și tuburilor și unui mediu alcalin, deja după 1-2 minute după actul sexual, ajung în uter, iar după 2-3 ore - secțiunile de capăt ale trompelor uterine, unde se contopesc de obicei cu oul. Există fertilizare monospermică (un spermatozoid intră în ovul) și polispermă (două sau mai mulți spermatozoizi pătrund în ovul, dar numai un nucleu spermatozoid se contopește cu nucleul ovulului). Conservarea activității spermatozoizilor în timpul trecerii lor în tractul genital al unei femei este facilitată de mediul ușor alcalin al canalului cervical al uterului, umplut cu un dop mucos. În timpul orgasmului în timpul actului sexual, dopul mucos de la canalul cervical este parțial împins în afară, apoi retras în el și, astfel, contribuie la o intrare mai rapidă a spermatozoizilor din vagin (unde o femeie sănătoasă are în mod normal un mediu ușor acid) într-un mediu mai acid. mediu favorabil al colului uterin și al cavității uterine. Trecerea spermatozoizilor prin dopul mucos al canalului cervical este facilitată și de creșterea bruscă a permeabilității mucusului în zilele ovulației. În zilele rămase ale ciclului menstrual, dopul mucos are o permeabilitate semnificativ mai scăzută pentru spermatozoizi.
Mulți spermatozoizi localizați în tractul genital al unei femei își pot păstra capacitatea de a fertiliza timp de 48-72 de ore (uneori chiar și până la 4-5 zile). Un ou ovulat rămâne viabil pentru aproximativ 24 de ore. Având în vedere acest lucru, cel mai favorabil moment pentru fertilizare este perioada de ruptură a unui folicul matur, urmată de nașterea unui ovul, precum și a 2-3-a zi după ovulație. Femeile care folosesc o metodă fiziologică de contracepție ar trebui să fie conștiente de faptul că momentul ovulației poate fluctua, iar viabilitatea ovulului și a spermei poate fi semnificativ mai lungă. La scurt timp după fertilizare, începe clivajul zigotului și formarea embrionilor.
zigotZigotul (zigotul grecesc pereche) este o celulă diploidă (conținând un set complet dublu de cromozomi) rezultată din fertilizare (fuziunea unui ovul și a unui spermatozoid). Zigotul este o celulă totipotentă (adică capabilă să producă orice altă celulă). Termenul a fost introdus de botanistul german E. Strasburger.
La om, prima diviziune mitotică a zigotului are loc la aproximativ 30 de ore după fertilizare, ceea ce se datorează proceselor complexe de pregătire pentru primul act de zdrobire. Celulele formate ca urmare a zdrobirii zigotului se numesc blastomeri. Primele diviziuni ale zigotului se numesc „zdrobire” deoarece celula este zdrobită: după fiecare diviziune, celulele fiice devin din ce în ce mai mici și nu există nicio etapă de creștere a celulelor între diviziuni.
Dezvoltarea zigotului Zigotul fie începe să se dezvolte imediat după fertilizare, fie își îmbracă o coajă densă și se transformă de ceva timp într-un spor de repaus (numit adesea zigospor) - tipic multor ciuperci și alge.
DespărțireaPerioada de dezvoltare embrionară a unui animal multicelular începe cu fragmentarea zigotului și se termină cu nașterea unui nou individ. Procesul de clivaj constă într-o serie de diviziuni mitotice succesive ale zigotului. Cele două celule formate ca urmare a unei noi diviziuni a zigotului și toate generațiile ulterioare de celule în acest stadiu se numesc blastomeri. În timpul zdrobirii, o diviziune urmează alteia, iar blastomerele rezultate nu cresc, drept urmare fiecare nouă generație de blastomere este reprezentată de celule mai mici. Această caracteristică a diviziunilor celulare în timpul dezvoltării unui ovul fecundat a determinat apariția unui termen figurat - zdrobirea zigotului.
La diferite specii de animale, ouăle diferă prin cantitatea și natura distribuției nutrienților de rezervă (gălbenuș) în citoplasmă. Acest lucru determină în mare măsură natura fragmentării ulterioare a zigotului. Cu o cantitate mică și o distribuție uniformă a gălbenușului în citoplasmă, întreaga masă a zigotului se împarte odată cu formarea de blastomere identice - zdrobire uniformă completă (de exemplu, la mamifere). Când gălbenușul se acumulează predominant la unul dintre polii zigotului, are loc o fragmentare neuniformă - se formează blastomeri care diferă ca mărime: macromere și micromere mai mari (de exemplu, la amfibieni). Dacă oul este foarte bogat în gălbenuș, atunci partea lui, fără gălbenuș, este zdrobită. Deci, la reptile, păsări, doar secțiunea în formă de disc a zigotului de la unul dintre poli, unde se află nucleul, suferă strivire - strivire incompletă, discoidală. În cele din urmă, la insecte, doar stratul de suprafață al citoplasmei zigotului este implicat în procesul de zdrobire - zdrobire incompletă, superficială.
Ca urmare a zdrobirii (atunci când numărul de blastomere divizate atinge un număr semnificativ), se formează o blastula. Într-un caz tipic (de exemplu, în lancetă), blastula este o bilă goală, al cărei perete este format dintr-un singur strat de celule (blastoderm). Cavitatea blastulei - blastocoel, denumită altfel cavitatea primară a corpului, este umplută cu lichid. La amfibieni, blastula are o cavitate foarte mică, iar la unele animale (cum ar fi artropodele), blastocelul poate fi complet absent.
gastrulatieÎn următoarea etapă a perioadei embrionare are loc procesul de formare a gastrulei - gastrulația. La multe animale, formarea gastrulei are loc prin invaginare, adică. proeminențe ale blastodermei la unul dintre polii blastulei (cu reproducere intensivă a celulelor în această zonă). Ca rezultat, se formează un embrion cu două straturi, în formă de bol. Stratul exterior al celulelor este ectodermul, iar stratul interior este endodermul. Cavitatea internă care apare atunci când iese peretele blastulei, intestinul primar, comunică cu mediul extern printr-o deschidere - gura primară (blastopor). Există și alte tipuri de gastrulație. De exemplu, la unele celenterate, endodermul gastrulei este format prin imigrare, i.e. „evacuarea” unei părți a celulelor blastodermice în cavitatea blastulei și reproducerea lor ulterioară. Gura primară se formează prin ruperea peretelui gastrulei. Cu strivire neuniformă (la unii viermi, moluște), gastrula se formează ca urmare a murdării macromerelor cu micromere și a formării endodermului datorită primului. Adesea sunt combinate diferite metode de gastrulare.
La toate animalele (cu excepția bureților și celenteratelor - animale cu două straturi), etapa de gastrulație se termină cu formarea unui alt strat de celule - mezodermul. Acest „strat celular se formează între ento- și ectoderm. Există două moduri cunoscute de așezare a mezodermului. În anelide, de exemplu, în regiunea blastoporă a gastrulei, sunt izolate două celule mari (teloblaste). Reproducând, ele dau se ridică la două dungi mezodermice, dintre care (parțial din cauza divergenței celulelor, parțial ca urmare a distrugerii unei părți a celulelor din interiorul benzilor mezodermice), se formează saci celomici - metoda teloblastică de așezare a mezodermului.În enterocelous metoda (echinoderma, lanceta, vertebrate), ca urmare a proeminenței peretelui intestinului primar, se formează buzunare laterale, care apoi se separă și devin celomice În ambele cazuri de formare a mezodermului, sacii celomici cresc și umplu cavitatea primară a corpului. Stratul mezodermic de celule care captuseste interiorul cavitatii corpului formeaza epiteliul peritoneal.Cavitatea care l-a inlocuit astfel pe cea primara se numeste cavitatea secundara a corpului, sau celom.In cazul metodei teloblastice de inchidere Mezodermul blastopor se dezvoltă în deschiderea orală a unui animal adult. Astfel de organisme sunt numite protostome. În deuterostomi (cu metoda enteroceloasă de depunere a mezodermului), blastoporul crește excesiv sau se transformă într-un anus, iar gura unui adult apare a doua oară, prin proeminența ectodermului.
Formarea a trei straturi germinale (ecto-, ento- si mezoderm) completeaza etapa de gastrulatie, iar din acest moment incep procesele de histo- si organogeneza. Ca rezultat al diferențierii celulare a celor trei straturi germinale, se formează diverse țesuturi și organe ale organismului în curs de dezvoltare. Încă de la sfârșitul secolului trecut (în mare parte datorită studiilor lui I. I. Mechnikov și A. O. Kovalevsky) s-a stabilit că la diferite specii de animale aceleași straturi germinale dau naștere acelorași organe și țesuturi. Din ectoderm se formează epiderma cu toate structurile derivate și sistemul nervos. Datorită endodermului se formează tractul digestiv și organele asociate (ficat, pancreas, plămâni etc.). Mezodermul formează scheletul, sistemul vascular, aparatul excretor, gonadele. Deși astăzi straturile germinale nu sunt considerate strict specializate, cu toate acestea, omologia lor în marea majoritate a speciilor de animale este evidentă, ceea ce indică unitatea originii regnului animal.
În timpul perioadei embrionare, există o creștere a ratei de creștere și diferențiere în organismele în curs de dezvoltare. Dacă creșterea nu are loc în procesul de zdrobire și blastula (prin masa sa) poate fi semnificativ inferioară zigotului, atunci, pornind de la procesul de gastrulare, masa embrionului crește rapid (datorită reproducerii celulare intensive). Procesele de diferențiere celulară încep în cea mai timpurie etapă a embriogenezei - zdrobirea și stau la baza diferențierii primare a țesuturilor - apariția a trei straturi germinale (țesuturi embrionare). Dezvoltarea ulterioară a embrionului este însoțită de un proces din ce în ce mai mare de diferențiere a țesuturilor și organelor. Ca urmare a perioadei embrionare de dezvoltare, se formează un organism care este capabil să existe independent (mai mult sau mai puțin) în mediul extern. Un nou individ se naște fie ca urmare a clocirii dintr-un ou (la animalele ovipare), fie ca urmare a ieșirii din corpul mamei (la vivipare).
Histo - și organogenezăHisto - și organogeneza embrionului sunt efectuate ca urmare a reproducerii, migrației, diferențierii celulelor, componentelor sale, stabilirii de contacte intercelulare și moartea unor celule. Din a 317-a până în a 20-a zi continuă perioada presomitică din a 20-a zi în care începe perioada somită de dezvoltare. În a 20-a zi de embriogeneză, prin formarea pliurilor trunchiului (cefalocaudale și laterale), embrionul însuși este separat de organele extraembrionare, precum și forma sa plată este schimbată în una cilindrică. În același timp, părțile dorsale ale mezodermului embrionului sunt împărțite în segmente separate situate pe ambele părți ale coardei - somite. În a 21-a zi, în corpul embrionului sunt 2-3 perechi de somiți. Somiții încep să se formeze din perechea III, perechile I și II apar ceva mai târziu. Numărul de somiți crește treptat: în a 23-a zi de dezvoltare sunt 10 perechi de somiți, în a 25-a - 14 perechi, în a 27-a - 25 de perechi, la sfârșitul celei de-a cincea săptămâni numărul de somiți din embrion ajunge la 43. -44 de perechi. Pe baza calculului numărului de somiți, este posibil să se determine aproximativ momentul de dezvoltare (vârsta somitică) a embrionului.
Din partea exterioară a fiecărui somit iese un dermatom, din interior - un sclerot, din mijloc - un miot. Dermatomul devine sursa dermului pielii, sclerotul devine sursa cartilajului și a țesutului osos, iar miotomul devine sursa mușchilor scheletici ai părții dorsale a embrionului. Secțiunile ventrale ale mezodermului - splanchnotom - nu sunt segmentate, ci sunt împărțite în foi viscerale și parietale, din care se dezvoltă membranele seroase ale organelor interne, țesutul muscular al inimii și cortexul suprarenal. Din mezenchimul splanhnotomului se formează vasele de sânge, celulele sanguine, țesutul muscular conjunctiv și neted al embrionului. Secțiunea mezodermului care leagă somiții cu splanchnotomul este împărțită în picioare segmentate - nefrogonotul, care servesc ca sursă pentru dezvoltarea rinichilor și gonadelor, precum și a canalelor paramezonefrice. Dintre acestea din urmă se formează epiteliul uterului și oviductul.
În procesul de diferențiere a ectodermului germinal se formează tubul neural, crestele neurale, placodele, ectodermul cutanat și placa precordală. Procesul de formare a tubului neural se numește neurulatie. Constă în formarea unei depresiuni sub formă de fante pe suprafața ectodermului; marginile îngroșate ale acestei depresiuni (pliuri neuronale) fuzionează pentru a forma tubul neural. Veziculele cerebrale se formează din partea craniană a tubului neural, care este rudimentul creierului. Pe ambele părți ale tubului neural (între acesta din urmă și ectodermul pielii), sunt separate grupuri de celule din care se formează crestele neurale. Celulele crestei neurale sunt capabile sa migreze. Celulele care migrează în direcția dermatomului dau naștere la celule pigmentare - melanocite; celulele crestei neurale care migreaza spre cavitatea abdominala dau nastere la ganglionii simpatici si parasimpatici, medula suprarenala. Din celulele crestelor neurale, care nu au migrat, se formează plăci ganglionare, din care se dezvoltă ganglionii nervilor autonomi spinali și periferici. Din placode se formează ganglionii capului și celulele nervoase ale organului auzului și echilibrului.
Antropologie și concepte de biologie Kurchanov Nikolai Anatolievici
Zigot și stadii de clivaj
Zigot și stadii de clivaj
zigot, format ca urmare a fuziunii gameților, este o etapă unicelulară în dezvoltarea unui organism multicelular. Deși durata acestei etape este de obicei scurtă, este posibil să se urmărească modificările citomorfologice și biochimice care apar în ea. Aceste modificări joacă un rol important în procesele ulterioare de embriogeneză. La un număr de animale, deja în zigot, sinteza proteinelor începe pe ARNm format în timpul oogenezei.
Despărțirea este un proces de diviziuni mitotice succesive, adesea inegale. Celulele care se formează în timpul clivajului sunt numite blastomeri. Zdrobirea se termină cu formarea blastula, având de obicei o cavitate internă - blastocoel. O trăsătură caracteristică a perioadei de zdrobire este lipsa creșterii. Deși embrionul în stadiul de blastulă poate consta din sute de celule, dimensiunea lui nu depășește dimensiunea zigotului.
În funcție de tipul de ouă, există mai multe tipuri de zdrobire și blastula. Există două tipuri principale de ouă.
Ouă homomolecite - au un nucleu situat in centru si un galbenus distribuit uniform in citoplasma.
Ouă telocitetale - au o polaritate pronunțată, o aranjare excentrică a nucleului și un gălbenuș distribuit neuniform în citoplasmă.
Ouăle homomolecite dau naștere de obicei blastule cu blastomeri identici: celoblastula(cu cavitate) sau morulu(fără cavitate). Ouăle telolecitale dau naștere blastulelor cu blastomere inegale: amfiblastula(zdrobire completă) sau discoblastula(zdrobire parțială). La mamifere, ca urmare a zdrobirii, se formează o morula, dar apoi, în timpul diviziunii asincrone complete a blastomerelor, are loc o etapă suplimentară - vezicule germinale, sau blastocist.
Acest text este o piesă introductivă. Din cartea Biologie [Un ghid complet pentru pregătirea pentru examen] autor Lerner Georgy Isaakovich Din cartea Fundamentals of Psychophysiology autor Alexandrov Yuri Din cartea Teoria nutriției adecvate și trofologie [tabele în text] autor1.2. Etapele fixării memoriei Ipoteza a două urme care se dezvoltă secvenţial. Conform ipotezei, formarea engramei se realizează în două etape: prima se caracterizează printr-o formă instabilă a urmei și există pentru o perioadă scurtă. Aceasta este scena
Din cartea Teoria nutriției adecvate și trofologie [tabele cu imagini] autor Ugolev Alexandru Mihailovici3. ETAPELE SOMMNULUI NREM ȘI ALE SOMNULUI REM Principalele date obținute de-a lungul anilor a numeroase și variate studii despre somn sunt următoarele. Somnul nu este o pauză în activitatea creierului, este doar o stare diferită. În timpul somnului, creierul trece prin mai multe faze diferite,
Din cartea Antropologie și concepte de biologie autor Kurchanov Nikolai Anatolievici Din cartea autorului9.2. Originea și etapele timpurii ale dezvoltării vieții Teoria modernă a originii vieții este teoria apariției structurilor care, dezvoltându-se și devenind mai complexe, au dobândit anumite proprietăți funcționale. Aparent, aceste proprietăți ar fi putut să apară deja în stadiile incipiente.
Din cartea autoruluiEtapele fotosintezei În procesul fotosintezei se disting două etape - lumina și întuneric.În timpul fazei luminoase a fotosintezei, energia Soarelui este folosită pentru a sintetiza ATP și purtători de electroni de înaltă energie. Energie luminoasă absorbită de orice moleculă
