Нервная ткань (при участии ряда других тканей) формирует нервную систему, обеспечивающую регуляцию всех жизненных процессов в организме и его взаимодействие с внешней средой.
Анатомически нервную систему делят на центральную и периферическую. К центральной относят головной и спинной мозг, периферическая объединяет нервные узлы, нервы и нервные окончания.
Нервная система развивается из нервной трубки и ганглиозной пластинки. Из краниальной части нервной трубки дифференцируются головной мозг и органы чувств. Из туловищного отдела нервной трубки - спинной мозг, из ганглиозной пластинки формируются спинномозговые и вегетативные узлы и хромаффинная ткань организма.
Нервные узлы (ганглии)
Нервные узлы, или ганглии, это скопления нейронов вне центральной нервной системы. Выделяют чувствительные и вегетативные нервные узлы.
Чувствительные нервные узлы лежат по ходу задних корешков спинного мозга и по ходу черепно-мозговых нервов. Афферентные нейроны в спиральном и вестибулярном ганглии являются биполярными, в остальных чувствительных ганглиях - псевдоуниполярными.
Спинномозговой узел (спинальный ганглий)
Спинномозговой узел имеет веретеновидную форму, окружен капсулой из плотной соединительной ткани. От капсулы в паренхиму узла проникают тонкие прослойки соединительной ткани, в которой расположены кровеносные сосуды.
Нейроны спинномозгового узла характеризуются крупным сферическим телом и светлым ядром с хорошо заметным ядрышком. Клетки располагаются группами, преимущественно по периферии органа. Центр спинномозгового узла состоит главным образом из отростков нейронов и тонких прослоек эндоневрия, несущих сосуды. Дендриты нервных клеток идут в составе чувствительной части смешанных спинномозговых нервов на периферию и заканчиваются там рецепторами. Аксоны в совокупности образуют задние корешки, несущие нервные импульсы в спинной мозг или продолговатый мозг.
В спинномозговых узлах высших позвоночных животных и человека биполярные нейроны в процессе созревания становятся псевдоуниполярными. От тела псевдоуниполярного нейрона отходит один отросток, который многократно обвивает клетку и часто образует клубок. Этот отросток разделяется Т-образно на афферентную (дендритную) и эфферентную (аксональную) ветви.
Дендриты и аксоны клеток в узле и за его пределами покрыты миелиновыми оболочками из нейролеммоцитов. Тело каждой нервной клетки в спинномозговом узле окружено слоем уплощенных клеток олигодендроглии, которые здесь называются мантийными глиоцитами, или глиоцитами ганглия, или же клетками-сателлитами. Они расположены вокруг тела нейрона и имеют мелкие округлые ядра. Снаружи глиальная оболочка нейрона покрыта тонковолокнистой соединительнотканной оболочкой. Клетки этой оболочки отличаются овальной формой ядер.
Нейроны спинномозговых узлов содержат такие нейромедиаторы, как ацетилхолин, глутаминовая кислота, вещество P.
Автономные (вегетативные) узлы
Вегетативные нервные узлы располагаются:
вдоль позвоночника (паравертебральные ганглии);
впереди от позвоночника (превертебральные ганглии);
в стенке органов - сердца, бронхов, пищеварительного тракта, мочевого пузыря (интрамуральные ганглии);
вблизи поверхности этих органов.
К вегетативным узлам подходят миелиновые преганглионарные волокна, содержащие отростки нейронов центральной нервной системы.
По функциональному признаку и локализации вегетативные нервные узлы разделяют на симпатические и парасимпатические.
Большинство внутренних органов имеет двойную вегетативную иннервацию, т.е. получает постганглионарные волокна от клеток, расположенных как в симпатических, так и в парасимпатических узлах. Реакции, опосредуемые их нейронами, часто имеют противоположную направленность (так, например, симпатическая стимуляция усиливает сердечную деятельность, а парасимпатическая ее тормозит).
Общий план строения вегетативных узлов сходен. Снаружи узел покрыт тонкой соединительнотканной капсулой. Вегетативные узлы содержат мультиполярные нейроны, которые характеризуются неправильной формой, эксцентрично расположенным ядром. Часто встречаются многоядерные и полиплоидные нейроны.
Каждый нейрон и его отростки окружены оболочкой из глиальных клеток-сателлитов - мантийных глиоцитов. Наружная поверхность глиальной оболочки покрыта базальной мембраной, кнаружи от которой расположена тонкая соединительнотканная оболочка.
Интрамуральные нервные узлы внутренних органов и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена иногда выделяются в самостоятельный метасимпатический отдел вегетативной нервной системы.
В интрамуральных узлах русским гистологом Догелем А.С. описаны нейроны трех типов:
длинноаксонные эфферентные клетки I типа;
равноотросчатые афферентные клетки II типа;
ассоциативные клетки III типа.
Длинноаксонные эфферентные нейроны (клетки Догеля I типа) - многочисленные и крупные нейроны с короткими дендритами и длинным аксоном, который направляется за пределы узла к рабочему органу, где образует двигательные или секреторные окончания.
Равноотросчатые афферентные нейроны (клетки Догеля II типа) имеют длинные дендриты и аксон, уходящий за пределы данного узла в соседние. Эти клетки входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС.
Ассоциативные нейроны (клетки Догеля III типа) - это местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типа.
Нейроны вегетативных нервных ганглиев, как и спинномозговых узлов, имеют эктодермальное происхождение и развиваются из клеток нервного гребня.
Периферические нервы
Нервы, или нервные стволы, связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами, или же с нервными узлами. Нервы образованы пучками нервных волокон, которые объединены соединительнотканными оболочками.
Большинство нервов - смешанные, т.е. включают афферентные и эфферентные нервные волокна.
Пучки нервных волокон содержат как миелиновые, так и безмиелиновые волокна. Диаметр волокон и соотношение между миелиновыми и безмиелиновыми нервными волокнами в различных нервах неодинаковы.
На поперечном срезе нерва видны сечения осевых цилиндров нервных волокон и одевающие их глиальные оболочки. В некоторых нервах встречаются одиночные нервные клетки и мелкие ганглии.
Между нервными волокнами в составе нервного пучка располагаются тонкие прослойки рыхлой волокнистой соединительной ткани - эндоневрий. В нем мало клеток, преобладают ретикулярные волокна, проходят мелкие кровеносные сосуды.
Отдельные пучки нервных волокон окружены периневрием. Периневрий состоит из чередующихся слоев плотно расположенных клеток и тонких коллагеновых волокон, ориентированных вдоль нерва.
Наружная оболочка нервного ствола - эпиневрий - представляет собой плотную волокнистую соединительную ткань, богатую фибробластами, макрофагами и жировыми клетками. Содержит кровеносные и лимфатические сосуды, чувствительные нервные окончания
Представляет собой систему тканей и органов, построенных из нервной ткани. В ней выделяют:
Центральный отдел: головной и спинной мозг
Периферический отдел: автономные и чувствительные ганглии, периферические нервы, нервные окончания.
Существует также подразделение на:
Соматический (анимальный, цереброспинальный) отдел;
Вегетативный (автономный) отдел: симпатическая и парасимпатическая части.
Нервную систему формируют следующие эмбриональные источники: нервная трубка, нервный гребень (ганглиозная пластинка) и эмбриональные плакоды. Тканевые элементы оболочек являются мезенхимными производными. На стадии замыкания нейропоров передний конец трубки значительно расширяется, боковые стенки утолщаются, образуя зачатки трех мозговых пузырей. Лежащий краниально пузырь образует передний мозг, средний пузырь - средний мозг, а из третьего пузыря, который переходит в закладку спинного мозга, развивается задний (ромбовидный) мозг. Вскоре после этого нервная трубка изгибается почти под прямым углом, и посредством борозд-сужений первый пузырь разделяется на конечный и промежуточный отделы, а третий мозговой пузырь - на продолговатый и задний отделы мозга. Производные среднего и заднего мозговых пузырей образуют ствол мозга и являются древними образованиями; в них сохраняется сегментарный принцип строения, который исчезает в производных промежуточного и конечного мозга. В последних концентрируются интегративные функции. Так формируются пять отделов мозга: конечный и промежуточный мозг, средний, продолговатый и задний мозг (у человека это происходит примерно в конце 4-й нед эмбрионального развития). Конечный мозг формирует два полушария большого мозга.
В эмбриональном гисто- и органогенезе нервной системы развитие разных отделов мозга происходит с разной скоростью (гетерохронно). Раньше формируются каудальные отделы центральной нервной системы (спинной мозг, ствол мозга); время окончательного формирования структур головного мозга сильно варьирует. В ряде отделов головного мозга это происходит после рождения (мозжечок, гиппокамп, обонятельная луковица); в каждом отделе мозга существуют пространственно-временные градиенты формирования нейронных популяций, которые образуют уникальную структуру нервного центра.
Спинной мозг представляет собой часть центральной нервной системы, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегмен-тарность. В боковых отделах нервной трубки быстро возрастает масса клеток, тогда как дорсальная и вентральная ее части не увеличиваются в объеме и сохраняют эпендимный характер. Утолщенные боковые стенки нервной трубки делятся продольной бороздой на дорсальную - крыльную, и вентральную - основную пластинку. На этой стадии развития в боковых стенках нервной трубки можно выделить три зоны: эпендиму, выстилающую центральный канал, промежуточную (плащевой слой) и маргинальную (краевую вуаль). Из плащевого слоя в дальнейшем развивается серое вещество спинного мозга, а из краевой вуали - его белое вещество. Нейробласты передних столбов дифференцируются в мотонейроны (двигательные нейроны) ядер передних рогов. Их аксоны выходят из спинного мозга и образуют передние корешки спинномозговых нервов. В задних столбах и промежуточной зоне развиваются различные ядра вставочных (ассоциативных) клеток. Их аксоны, поступая в белое вещество спинного мозга, входят в состав различных проводящих пучков. В задние рога входят центральные отростки чувствительных нейронов спинномозговых узлов.
Одновременно с развитием спинного мозга закладываются спинномозговые и периферические узлы автономной нервной системы. Исходным материалом для них служат стволовые клеточные элементы нервного гребня, которые путем дивергентной дифференцировки развиваются в нейробла-стическом и глиобластическом направлениях. Часть клеток нервного гребня мигрирует на периферию в места локализации узлов автономной нервной системы, параганглиев, нейроэндокринных клеток APUD-серии и хромаффинной ткани.
Периферическая нервная система.
Периферическая нервная система объединяет периферические нервные узлы, стволы и окончания.
Нервные ганглии (узлы) – структуры, образованные скоплениями нейронов вне ЦНС, - разделяются на чувствительные и автономные (вегетативные). Чувствительные ганглии содержат псевдоуниполярные или биполярные (в спиральном и вестибулярном ганглиях) афферентные нейроны и располагаются преимущественно по ходу задних корешков спинного мозга (чувствительные узлы спинномозговых нервов) и некоторых черепно-мозговых нервов. Чувствительные ганглии спинномозговых нервов имеют веретеновидную форму и покрыты капсулой из плотной волокнистой соединительной ткани. По периферии ганглия находятся плотные скопления тел псевдоуниполярных нейронов, а центральная часть занята их отростками и расположенными между ними тонкими прослойками эндоневрия, несущими сосуды. Автономные нервные ганглии образованы скоплениями мультиполярных нейронов, на которых многочисленные синапсы образуют преганглионарные волокна – отростки нейронов, чьи тела лежат в ЦНС.
Нерв. Строение и регенерация. Спинномозговые ганглии. Морфофункциональная характеристика.
Нервы (нервные стволы) связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами. Они образованы пучками миелиновых и безмиелиновых волокон, которые объединены соединительнотканными компонентамии (оболочками): эндоневрием, периневрием и эпиневрием. Большинство нервов являются смешанными, т.е. включают афферентные и эфферентные волокна.
Эндоневрий – тонкие прослойки рыхлой волокнистой соединительной ткани с мелкими кровеносными сосудами, окружающие отдельные нервные волокна и связывающие их в единый пучок. Периневрий – оболочка, покрывающая каждый пучок нервных волокон снаружи и отдающая перегородки вглубь пучка. Он имеет пластинчатое строение и образов концентрическими пластами уплощённых фиброблатстоподобных клеток, связанных плотными и щелевыми соединениями. Между слоями клеток в пространствах, заполненных жидкостью, располагаются компоненты базальной мембраны и продольно ориентированные коллагеновые волокна. Эпиневрий – наружная оболочка нерва, связывающая воедино пучки нервных волокон. Он состоит из плотной волокнистой соединительной ткани, содержащей жировые клетки, кровеносные и лимфатические сосуды.
Спинной мозг. Морфофункциональная характеристика. Развитие. Строение серого и белого вещества. Нейронный состав.
Спинной мозг состоит из двух симметричных половин, отграниченных друг от друга спереди глубокой серединной щелью, а сзади – соединительнотканной перегородкой. Внутренняя часть органа темнее - это его серое вещество. На периферии спинного мозга располагается более светлое белое вещество. Серое вещество спинного мозга состоит из тел нейронов, безмиелиновых и тонких миелиновых волокон и нейроглии. Основной составной частью серого вещества, отличающей его от белого, являются мультиполярные нейроны. Выступы серого вещества принято называть рогами. Различают передние, или вентральные, задние, или дорсальные, и боковые, или латеральные, рога. В процессе развития спинного мозга из нервной трубки образуются нейроны, группирующиеся в 10 слоях, или в пластинах. Для человека характерна
следующая архитектоникауказанных пластин: I-V пластины соответствуют задним рогам, VI-VII пластины - промежуточной зоне, VIII-IX пластины - передним рогам, X пластина - зона околоцентрального канала. Серое вещество мозга состоит из мультиполярных нейронов трех типов. Первый тип нейронов является филогенетически более древним и характеризуется немногочисленными длинными, прямыми и слабо ветвящимися дендритами (изодендритический тип). Второй тип нейронов имеет большое число сильно ветвящихся дендритов, которые переплетаются, образуя «клубки» (идиодендритический тип). Третий тип нейронов по степени развития дендритов занимает промежуточное положение между первым и вторым типами. Белое вещество спинного мозга представляет собой совокупность продольно ориентированных преимущественно миелиновых волокон. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга
Головной мозг. Источники развития. Общая морфофункциональная характеристика больших полушарий. Нейронная организация больших полушарий. Цито- и миелоархитектоника коры больших полушарий головного мозга. Возрастные изменения коры.
В головном мозге различают серое и белое вещество, но распределение этих двух составных частей здесь значительно сложнее, чем в спинном мозге. Большая часть серого вещества головного мозга располагается на поверхности большого мозга и в мозжечке, образуя их кору. Меньшая часть образует многочисленные ядра ствола мозга.
Строение . Кора большого мозга представлена слоем серого вещества. Наиболее сильно развита она в передней центральной извилине. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга.. Различные участки ее, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток (цитоархитектоника), расположения волокон (миелоархитектоника) и функциональным значением, называются полями. Они представляют собой места высшего анализа и синтеза нервных импульсов. Резко очерченные
границы между ними отсутствуют. Для коры характерно расположение клеток и волокон слоями. Развитие коры больших полушарий (неокортекса) человека в эмбриогенезе происходит из вентрикулярной герминативной зоны конечного мозга, где расположены малоспециализированные пролиферирующие клетки. Из этих клеток дифференцируются нейроциты неокортекса. При этом клетки утрачивают способность к делению и мигрируют в формирующуюся корковую пластинку. Вначале в корковую пластинку поступают нейроциты будущих I и VI слоев, т.е. наиболее поверхностного и глубокого слоев коры. Затем в нее встраиваются в направлении изнутри и кнаружи последовательно нейроны V, IV, III и II слоев. Этот процесс осуществляется за счет образования клеток в небольших участках вентрикулярной зоны в различные периоды эмбриогенеза (гетерохрон-но). В каждом из этих участков образуются группы нейронов, последовательно выстраивающихся вдоль одного или нескольких волокон
радиальной глии в виде колонки.
Цитоархитектоника коры большого мозга. Мультиполярные нейроны коры весьма разнообразны по форме. Среди них можно выделить пирамидные, звездчатые, веретенообразные, паукообразные и горизонтальные нейроны. Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоев: I - молекулярный, II - наружный зернистый, III - nuрамидных нейронов, IV - внутренний зернистый, V - ганглионарный, VI - слой полиморфных клеток. Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их нейриты проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Наружный зернистый слой образован мелкими нейронами, имеющими округлую, угловатую и пирамидальную форму, и звездчатыми нейроцитами. Дендриты этих клеток поднимаются в молекулярный слой. Нейриты или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя. Самый широкий слой коры большого мозга - пирамидный. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Нейрит пирамидной клетки всегда отходит от ее основания. Внутренний зернистый слой образован мелкими звездчатыми нейронами. В его состав входит большое количество горизонтальных волокон. Ганглионарный слой коры образован крупными пирамидами, причем область прецентральной извилины содержит гигантские пирамиды.
Слой полиморфных клеток образован нейронами различной формы.
Миелоархитектоника коры . Среди нервных волокон коры полушарий большого мозга можно выделить ассоциативные волокна, связывающие отдельные участки коры одного полушария, комиссуральные, соединяющие кору различных полушарий, и проекционные волокна, как афферентные, так и эфферентные, которые связывают кору с ядрами низших отделов центральной
нервной системы.
Возрастные изменения . На 1-м году жизни наблюдаются типизация формы пирамидных и звездчатых нейронов, их увеличение, развитие дендритных и аксонных арборизаций, внутриансамблевых связей по вертикали. К 3 годам в ансамблях выявляются «гнездные» группировки нейронов, более четко сформированные вертикальные дендритные пучки и пучки радиарных волокон. К 5-6 годам нарастает полиморфизм нейронов; усложняется система внутриансамблевых связей по горизонтали за счет роста в длину и разветвлений боковых и базальных дендритов пирамидных нейронов и развития боковых терминалей их апикальных дендритов. К 9-10 годам увеличиваются клеточные группировки, значительно усложняется структура короткоаксонных нейронов, и расширяется сеть аксонных коллатералей всех форм интернейронов. К 12-14 годам в ансамблях четко обозначаются специализированные формы пирамидных нейронов, все типы интернейронов достигают высокого уровня дифференцировки. К 18 годам ансамблевая организация коры по основным параметрам своей архитектоники достигает уровня таковой у взрослых.
Мозжечок. Строение и морфофункциональная характеристика. Нейронный состав коры мозжечка, глиоциты. Межнейронные связи.
Мозжечок . Представляет собой центральный орган равновесия и координации движений. Он связан со стволом мозга афферентными и эфферентными проводящими пучками, образующими в совокупности три пары ножек мозжечка. На поверхности мозжечка много извилин и бороздок, которые значительно увеличивают ее площадь. Борозды и извилины создают на разрезе
характерную для мозжечка картину «древа жизни». Основная масса серого вещества в мозжечке располагается на поверхности и образует его кору. Меньшая часть серого вещества лежит глубоко в белом веществе в виде центральных ядер. В центре каждой извилины имеется тонкая прослойка
белого вещества, покрытая слоем серого вещества - корой. В коре мозжечка различают три слоя: наружный - молекулярный, средний - ганглионарный слой, или слой грушевидных нейронов, и внутренний -зернистый. Ганглиозный слой содержит грушевидные нейроны. Они имеют нейриты, которые, покидая кору мозжечка, образуют начальное звено его эфферентных
тормозных путей. От грушевидного тела в молекулярный слой отходят 2-3 дендрита, которые пронизывают всю толщу молекулярного слоя. От основания тел этих клеток отходят нейриты, проходящие через зернистый слой коры мозжечка в белое вещество и заканчивающиеся на клетках ядер мозжечка. Молекулярный слой содержит два основных вида нейронов: кор-зинчатые и звездчатые. Корзинчатые нейроны находятся в нижней трети молекулярного слоя. Их тонкие длинные дендриты ветвятся преимущественно в плоскости, расположенной поперечно к извилине. Длинные нейриты клеток всегда идут поперек извилины и параллельно поверхности над грушевидными нейронами. Звездчатые нейроны лежат выше корзинчатых и эывают двух типов. Мелкие звездчатые нейроны снабжены тонкими короткими дендритами и слаборазветвленными нейритами, образующими синапсы. Крупные звездчатые нейроны имеют длинные и сильно разветвленные дендриты и нейриты. Зернистый слой. Первым типом клеток этого слоя можно считать зерновидные нейроны, или клетки-зерна. Клетка имеет 3-4 коротких дендрита,
заканчивающихся в этом же слое концевыми ветвлениями в виде лапки птицы. Нейриты клеток-зерен проходят в молекулярный слой и в нем делятся на две ветви, ориентированные параллельно поверхности коры вдоль извилин мозжечка. Вторым типом клеток зернистого слоя мозжечка являются тормозные большие звездчатые нейроны. Различают два вида таких клеток: с короткими и длинными нейритами. Нейроны с короткими нейритами лежат вблизи ганглионарного слоя. Их разветвленные дендриты распространяются в молекулярном слое и образуют синапсы с параллельными волокнами - аксонами клеток-зерен. Нейриты направляются в зернистый слой к клубочкам мозжечка и заканчиваются синапсами на концевых ветвлениях дендритов клеток-зерен.
Немногочисленные звездчатые нейроны с длинными нейритами имеют обильно ветвящиеся в зернистом слое дендриты и нейриты, выходящие в белое вещество. Третий тип клеток составляют веретеновидные горизонтальные клетки. Они имеют небольшое вытянутое тело, от которого в обе стороны отходят длинные горизонтальные дендриты, заканчивающиеся в ганглионарном и зернистом слоях. Нейриты же этих клеток дают коллатерали в зернистый слой и уходят в
белое вещество. Глиоциты . Кора мозжечка содержит различные глиальные элементы. В зернистом слое имеются волокнистые и протоплазматические астроциты. Ножки отростков волокнистых астроцитов образуют периваскулярные мембраны. Во всех слоях в мозжечке имеются олигодендроциты. Особенно богаты этими клетками зернистый слой и белое вещество мозжечка. В ганглионарном слое между грушевидными нейронами лежат глиальные клетки с темными ядрами. Отростки этих клеток направляются к поверхности коры и образуют глиальные волокна молекулярного слоя мозжечка. Межнейрональные связи . Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами - моховидными и так называемыми лазящими волокнами. Моховидные волокна идут в составе оливомозжечкового и мостомозжечкового путей и опосредованно через клетки-зерна оказывают на грушевидные клетки возбуждающее действие.
Лазящие волокна поступают в кору мозжечка, по-видимому, по спинно-мозжечковому и вестибуломозжечковому путям. Они пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами. Лазящие волокна передают возбуждение непосредственно грушевидным нейронам.
Автономная (вегетативная) нервная система. Общая морфофункциональная характеристика. Отделы. Строение экстрамуральных и интрамуральных ганглиев.
ВНС делится на симпатическую и парасимпатическую. Обе системы одновременно принимают участие в иннервации органов и оказывают на них противоположное влияние. Состоит из центральных отделов, представленных ядрами серого вещества головного и спинного мозга, и периферических: нервных стволов, узлов (ганглиев) и сплетений.
Интрамуральные ганглии и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена выделяют в самостоятельный метасимпатический отдел автономной НС. Выделяют нейроны трёх типов:
Длинноаксонные эфферентные нейроны (клетки I типа Догеля) с короткими дендритами и длинным аксоном, идущим за пределы узла к клеткам рабочего органа, на которых он образует двигательные или секреторные окончания.
Равноотросчатые афферентные нейроны (клетки II типа Догеля) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС.
Ассоциативные клетки (клетки III типа Догеля) – местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типов. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
Подробности
Ганглии представляют собой скопления мультиполярных (один аксон и несколько дендритов) нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия.
Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны . В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля 1-го типа), чувствительные (клетки Догеля П-го типа) и ассоциативные (клетки Догеля Ш-го типа) . Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки 1-го типа называют длинноаксонными нейронами. Нейроны П-го типа - равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки П-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля Ш-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.
Ацетилхолин
- никотиновые (блок кураре, гексаметоний), мускариновые (блок атропин) рецепторы. Активация рецепторов→генерирование ВПСП. Быстрый ВПСП (N-холиноцер)→открытие ионных каналов. Медленный ВПСП (М-холинорец)→подавление М-тока, обусловленного повышением К-проводимости.
Нейропептиды
– действуют как нейромодуляторы.
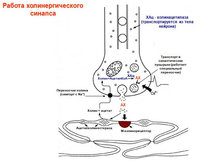
Энкефалины
, вещество Р, люлиберин, нейротензин, соматостатин – симп. ганглии (+Ach)
Катехоламины
(NA, дофамин) – нейромедиаторы мелких клеток с интенсивной флуоресценцией.
Нейропептид Y, соматостатин – симп. постганглионары.
Симпатические постганглионары: NA, АТФ, нейропептид У.
α1→инозотолтрифосфат, диацилглицерол. α2→активация G-белка, ↓цАМФ.
β→G-белок→AC→цАМФ
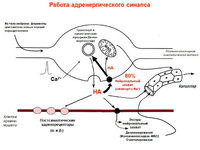
Исключения
: медиатор Ach, рецепторы мускариновые.
Парасимп. постганглионары: Ach, ВИП, NO, соматостатин, АТФ, опиоидные пептиды.
М1 (высокое сродство к пирензепину)-повышение секреции кислоты клетками желез желудка, М2 (низкое)- замедляют серд. ритм, секреция слезных и слюнные желез.
Разнообразное действие:
-Специфические втор. посредники: М2 может акт-ть IP3, а может инг-ть AC, уменьшая цАМФ.
-Действие на К и Са-каналы
-На эндотелии образуется NO→гуанилатциклаза→цГМФ→ цГМФ-зависимая протеинкиназа→расслабление глажких мышц.
Частная гистология.
Частная гистология – наука о микроскопическом строении и происхождении органов. Каждый орган состоит из 4 тканей.
Органы нервной системы.
По функциональному признаку
1. Соматическую нервную систему – участвует в иннервации тела человека и высшей нервной деятельности.
a. Центральный отдел :
i. Спинной мозг – ядра задних и передних рогов
ii. Головной мозг – кора мозжечка и больших полушарий
b. Периферический отдел :
i. Спинномозговые ганглии
ii. Черепномозговые ганглии
iii. Нервные стволы
2. Вегетативную нервную систему – обеспечивает работу внутренних органов, иннервирует гладкие миоциты и представляет секреторные нервы.
1) Симпатическая :
a. Центральный отдел :
i. Спинной мозг – ядра боковых рогов торако-люмбального отдела
ii. Головной мозг – гипоталамус
b. Периферический отдел :
i. Симпатические ганглии
ii. Нервные стволы
2) Парасимпатическая :
a. Центральный отдел :
i. Спинной мозг – ядра боковых рогов сакрального отдела
ii. Головной мозг – ядра ствола, гипоталамус
b. Периферический отдел :
i. Парасимпатические ганглии
ii. Нервные стволы
iii. Спинномозговые и черепно-мозговые ганглии
По анатомическому признаку органы нервной системы делятся на:
1. Периферическую нервную систему.
2. Центральную нервную систему.
Эмбриональные источники развития :
1. Нейроэктодерма (даёт начало паренхиме органов).
2. Мезенхима (даёт начало строме органов, совокупности вспомогательных структур, обеспечивающих функционирование паренхимы).
Органы нервной системы функционируют в относительной изоляции от окружающей среды, отделяясь от неё биологическими барьерами . Виды биологических барьеров :
1. Гематонейральный (отграничивает кровь от нейронов).
2. Ликворонейральный (отграничивает ликвор от нейронов).
3. Гематоликворный (отграничивает ликвор от крови).
Функции нервной системы:
1. Регуляция функций отдельных внутренних органов.
2. Интеграция внутренних органов в системы органов.
3. Обеспечение взаимосвязи организма с внешней средой.
4. Обеспечение высшей нервной деятельности.
Все функции основываются на принципе рефлекса . Материальной основой является рефлекторная дуга , состоящая из 3-х звеньев: афферентного , ассоциативного и эфферентного . Они распределены по отдельным органам нервной системы.
Органы периферической нервной системы :
1. Нервные стволы (нервы).
2. Нервные узлы (ганглии).
3. Нервные окончания.
Нервные стволы – это пучки нервных волокон, объединённые системой соединительно-тканных оболочек. Нервные стволы смешаны, т.е. в каждом есть миелиновые и амиелиновые волокна, в результате чего обслуживается соматическая и вегетативная нервные системы.
Строение нервного ствола :
1. Паренхима : безмиелиновые и миелиновые нервные волокна + микроганглии.
2. Строма : соединительно-тканные оболочки:
1) Периневрий (периневральные влагалища: РВНСТ + кровеносные сосуды + эпендимоглиоциты + цереброспинальная жидкость).
2) Эпиневрий (ПВНСТ + кровеносные сосуды).
3) Периневрий (отщепление от эпиневрия внутрь ствола).
4) Эндоневрий (РВНСТ + кровеносные сосуды).
В периневрии есть щелевидное пространство – щелевидное периневральное влагалище , которое заполнено ликвором (циркулирующей биологической жидкостью). Структурные компоненты стенок периневрального влагалища :
1. Низкопризматические эпендимоглиоциты.
2. Базальная мембрана.
3. Субэпендимальная пластинка.
4. Кровеносные сосуды.
Ликвор в периневральном влагалище может отсутствовать. В них иногда вводят анестезирующие вещества, антибиотики (т.к. по ним распространяется заболевание).
Функции нервных стволов :
1. Проводниковая (проводят нервный импульс).
2. Трофическая (питательная).
4. Являются начальным звеном в секреции и циркуляции цереброспинальной жидкости.
Регенерация нервных стволов :
1. Физиологическая регенерация (очень активное восстановление оболочек за счёт фибробластов).
2. Репаративная регенерация (восстанавливается тот участок нервного ствола, нервные волокна которого не утратили связи с перикарионом – они способны расти на 1мм/сутки; периферические отрезки нервных волокон не восстанавливаются).
Нервные узлы (ганглии) – группы или кооперации нейронов, вынесенные за пределы мозга. Нервные узлы «одеты» в капсулы.
Типы ганглиев:
1. Спинномозговые .
2. Черепномозговые .
3. Вегетативные .
Спинномозговые ганглии – утолщения на начальных отделах задних корешков спинного мозга; это скопление афферентных (чувствительных) нейронов (они являются первыми нейронами в цепи рефлекторной дуги).
Строение спинномозгового ганглия :
1. Строма :
1) наружная соединительно-тканная капсула, состоящая из 2 листков:
a. наружный листок (плотная соединительная ткань – продолжение эпиневрия спинномозгового нерва)
b. внутренний листок (многотканевой: РВНСТ, глиоциты; аналог периневрия спинномозгового нерва; есть расщепления, переходящие на внутриорганные перегородки, заполнены ликвором).
2) внутриорганные перегородки, отходящие от капсулы внутрь узла
b. кровеносные и лимфатические сосуды
c. нервные волокна
d. нервные окончания
3) собственные соединительно-тканные капсулы псевдоуниполярных нейронов
a. волокнистая соединительная ткань
b. однослойный плоский эпендимоглиальный эпителий
c. перинейрональное пространство с цереброспинальной жидкостью
2. Паренхима :
1) центральная часть (миелиновые нервные волокна – отростки псевдоуниполярных нейронов)
2) периферическая часть (псевдоуниполярные нейроны + мантийные глиоциты (олигодендроглиоциты)).
Функции спинномозгового ганглия:
1. Участие в рефлекторной деятельности (первые нейроны в цепи рефлекторной дуги).
2. Являются начальным звеном в обработке афферентной информации.
3. Барьерная функция (гематонейральный барьер).
4. Являются звеном в циркуляции ликвора.
Источники эмбрионального развития спинномозгового ганглия :
1. Ганглиозная пластинка (даёт начало элементам паренхимы органа).
2. Мезенхима (даёт начало элементам стромы органа).
Ганглии вегетативной нервной системы – расположены после спинного мозга, участвуют в создании вегетативных дуг.
Типы ганглиев вегетативной нервной системы :
1. Симпатические :
1) Паравертебральные;
2) Превертебральные;
2. Парасимпатические :
1) Внутриорганные (интрамуральные);
2) Околоорганные (параорганные);
3) Вегетативные узлы головы (по ходу черепно-мозговых нервов).
Строение ганглиев вегетативной нервной системы :
1. Строма : строение аналогично строме спинномозгового ганглия.
2.1. Паренхима симпатических ганглиев : нейроны, расположенные хаотично по всему ганглию + клетки-сателлиты + соединительно-тканная капсула.
1) крупные длинноаксонные мультиполярные эфферентные адренергические нейроны
2) мелкие равноотростчатые мультиполярные ассоциативные адренергические интенсивно флюоресцирующие (МИФ) – нейроны
3) преганглионарные миелиновые холинэргические волокна (аксоны нейронов боковых рогов спинного мозга)
4) постганглионарные безмиелиновые адренергические нервные волокна (аксоны крупных нейронов ганглиев)
5) внутриганглионарные безмиелиновые ассоциативные нервные волокна (аксоны МИФ – нейронов).
2.2. Паренхима парасимпатических ганглиев :
1) длинноаксонные мультиполярные эфферентные холинэргические нейроны (Догеля I типа).
2) длиннодендритные мультиполярные афферентные холинэргические нейроны (Догеля II типа): дендрит – к рецептору, аксон – к 1 и 3 типу.
3) равноотростчатые мультиполярные ассоциативные холинэргические нейроны (Догеля III типа).
4) преганглионарные миелиновые холинергические нервные волокна (аксоны боковых рогов спинного мозга).
5) постганглионарные безмиелиновые холинергические нервные волокна (аксоны нейронов Догеля I типа).
Функции ганглиев вегетативной нервной системы :
1. Симпатических :
1) Проведение импульсов к рабочим органам (2.1.1)
2) Распространение импульса в пределах ганглия (тормозной эффект) (2.1.2)
2. Парасимпатических :
1) Проведение импульса к рабочим органам (2.2.1)
2) Проведение импульса от интерорецепторов в пределах местных рефлекторных дуг (2.2.2)
3) распространение импульса в пределах или между ганглиями (2.2.3).
Источники эмбрионального развития ганглиев вегетативной нервной системы :
1. Ганглиозная пластинка (нейроны и нейроглия).
2. Мезенхима (соединительная ткань, сосуды).
Кроме парасимпатического и симпатического отделов, физиологи выделяют метасимпатический отдел вегетативной нервной системы. Под этим термином подразумевается комплекс микроганглионарных образований, расположенных в стенках вн>тренних органов, обладающих моторной активностью (сердце, кишечник, мочеточники и др.) и обеспечивающих их автономию. Функция нервных узлон заключается В передаче центральных (симпатических, парасим-патических) влияний к тканям, и, кроме того, они обеспечивают интеграцию информации, поступающей по местным рефлекторным дугам. Метасимпати-ческие структуры представляют собой самостоятельные образования, способные функционировать при полной децентрализии. Несколько (5—7) из относящихся к ним близлежащих узлов объединяются в единый функциональный модуль, основными единицами которого являются клетки-осцилляторы, обеспечи-вающие автономию системы, интернейроны, мотонейроны, чувствительные клетки. Отдельные функциональные модули составляют сплетение, благодаря которому, например, в кишке организуется перистальтическая волна.
Функции метасимпатического отдела вегетативной нервной системы на-прямую не зависят от деятельности симпатической или парасимпатической
нервных систем, но могут видоизменяться под их влиянием. Так, например, активация парасимпатического влияния усиливает перистальтику кишечника, а симпатического — ослабляет ее.
- Многочисленные небольшие скопления нервных клеток, входящие в состав обширных нервных сплетений в стенках внутренних органов (желудочно-кишечного тракта, сердца и др.), иногда относят к парасимпатическому отделу вегетативной нервной системы на том основании, что в морфологических работах легко обнаруживаются синаптические контакты между этими клетками и волокнами блуждающего нерва.
- Метасимпатическая нервная система, интрамуральные нервные сплетения обнаружены в сердце, всех полых органах, но глубже изучены на примере иннервации желудка и кишечника. В этих отделах желудочно-кишечного тракта интрагастральная и энтеральная нервная система представлена настолько обильно, что по числу нейронов (108 единиц) сопоставима со спинным мозгом. Это дает основание для образного названия ее «брюшной мозг».
- По ответам на длительный толчок деполяризующего тока все эн-теральные нейроны межмышечного сплетения можно разделить на два типа: первый - тип S и второй - тип АН. Нейроны типа S отвечают на это раздражение длительной серией спайков, а нейроны типа АН - лишь одним или двумя спайками, которые сопровождаются сильной и длительной (4-20 с) следовой гиперполяризацией, отсутствующей у типа S. Спайк у нейронов типа S обусловливается натриевой, а у нейронов типа АН - натриевой и кальциевой проводимостью мемб-раны.
- ПМ - продольная мышца, МС - межмышечное сплетение, К.М - круговая мышца, ПС - подслизистое сплетение, С - слизистая оболочка; указаны нейроны, содержащие или выде-ляющие ацетилхолин [А X}, серотонин (5-гидрокситриптамин (5-ГТ)) и различные пептиды (вызывающие возбуждающие (+) или тормозные МХР - мускариновые холинорецепторы, а-А Р - альфа-адренорецепторы.
Метасимпатическая нервная система (МНС) в целом состоит из нервных узлов и сплетений, расположенных в толще внутренних органов. МНС отличается от других отделов нервной системы рядом особенностей:
1. Иннервирует только внутренние органы, наделенные собственной двигательной активностью;
2. Не имеет непосредственных контактов с рефлекторными дугами соматической нервной системы; получает синаптические входы только от симпатической и парасимпатической систем;
3. Наряду с общими для всей вегетативной системы афферентными путями, имеет и собственное чувствительное звено;
4. Не проявляет влияний, противоположных действию других частей ВНС, что характерно для симпатической и парасимпатической систем;
5. Обладает значительно большей автономностью, чем другие части ВНС.
В желудочно-кишечном тракте выражены все основные характеристики строения и функционирования метасимпатической нервной системы, и, кроме того, именно в ЖКТ эта система изучена наиболее полно по сравнению со всеми остальными органами. Поэтому ЖКТ является наиболее подходящим объектом для ознакомления с МНС.
В состав ЖКТ входят самые разные эффекторные образования – гладкая мышечная ткань, эпителий слизистых оболочек, железы, кровеносные и лимфатические сосуды, элементы иммунной системы, эндокринные клетки. Регуляция и координация деятельности всех этих структур осуществляют местная энтеральная метасимпатическая нервная система, с участием симпатического и парасимпатического отделов вегетативной нервной системы и висцеральных афферентов, образованных нейронами спинномозговых узлов. Большинство простейших функций ЖКТ не нарушается при разрыве экстраорганных (парасимпатических и симпатических) нервных путей.
Клеточные тела большинства нейронов энтеральной метасимпатической нервной системы залегают в нервных сплетениях (в ганглиях и внутри нервных стволов).
У человека в стенках пищевода, желудка и кишечника находятся три связанных между собой сплетения: подсерозное, межмышечное (Ауэрбаха) и подслизистое (Мейснера). Подсерозное сплетение наиболее представлено на дне и в большой кривизне желудка и состоит из мелких, густо расположенных скоплений нейронов и нервных волокон. В кишечнике элементы этого сплетения сосредоточены главным образом под мышечными лентами ободочной кишки. Наиболее массивным из всех нервных сплетений ЖКТ является межмышечное , расположенное между циркулярным и продольным слоями мышечной оболочки. В стенке желудка это сплетение имеет вид многослойной сети, и его плотность возрастает от дна к пилорической части. В области привратника в составе сплетения имеется огромная масса узлов, которые образуют обширные клеточные поля. Крупные (до 60 нейронов), средние и мелкие (2-8 нейронов) узлы располагаются по ходу нервных стволиков и в местах их ветвлений. Число нейронов на 1 см2 достигает 2000. Сильно развито межмышечное сплетение и в стенке тонкой кишки. Здесь ганглии в основном мелкие, содержат 5-20 нейронов.
Подслизистое сплетение представляет собой узкопетлистую сеть нервных пучков и микроганглиев, содержащих 5-15 (редко до 30) нейронов. Оно имеет поверхностную и глубокую части. Ветви этого сплетения подходят к основаниям выводных протоков желез и образуют межжелезистое сплетение. Тонкие волокна оканчиваются на эпителиальных клетках. Строение подслизистого сплетения по длине пищеварительного тракта изменяется незначительно, только в пищеводе оно развито слабо. По данным сканирующей электронной микроскопии, поверхностное подслизистое сплетение во всех частях тонкой кишки располагается непосредственно под мышечным слоем слизистой оболочки, отдает в этот слой многочисленные пучки диаметром 1-20 мкм. Такими же пучками связаны и отдельные узлы, диаметр которых составляет 20-400 иногда до 800 мкм. Узлы покрыты сплошным слоем фибробластов и коллагеновых волокон, после удаления которых видны контуры нейронов, а на их поверхности различимы многочисленные тонкие отростки. Однако нейроны в целом не выявляются, так как они окружены отростками клеток глии.
Во все части межмышечного и подслизистого сплетений вступают стволики неорганных нервов (симпатических, парасимпатических) (Рис.10). Размеры нейронов и узлов, их количество в сплетениях сильно варьирует в различных частях ЖКТ. Так, у человека средних лет в нижней трети пищевода в межмышечном сплетении имеются крупные узлы, диаметром до 960 мкм, содержащие 50-60 (иногда до 85 нейронов), в то время как в узлах подслизистого сплетения пищевода содержится лишь по 10-15 нейронов.
В узлах энтеральной метасимпатической системы наряду с дифференцированными нейронами диаметром 30-58 мкм имеются мелкие малодифференцированные клетки.
Известный русский гистолог А.С. Догель в результате исследований нейронов интрамуральных узлов пищеварительного тракта выделил три типа клеток. (Рис.11) К I типу относятся клетки средних размеров с округлым перикарионом, хорошо выраженным, длинным аксоном и многочисленными (до 20) короткими дендритами с широким основанием. Они отличаются от других нейронов узла тинкториальными свойствами: слабо импрегнируются азотнокислым серебром, но хорошо окрашиваются метиленовой синью. На препаратах, импрегнированных серебром, имеют темное крупное ядро и светлую цитоплазму. Дендриты не выходят за пределы узла, сильно ветвятся, образую густое сплетение, вступают в много численные контакты с другими нейронами. Эти клетки являются эфферентными; их аксоны покидают узел и оканчиваются варикозными терминалями на пучках гладких миоцитов и железах. На клетках Догеля I типа заканчиваются преганглионарные парасимпатические волокна из дорзального ядра блуждающего нерва, а также симпатические преганглионары из промежуточнобокового ядра спинного мозга.
Рис. 11. Схема межнейронных связей энтеральной части МНС.
1 – чувствительный нейрон; 2 – интернейрон; 3 – эфферентный нейрон; 4 – постганглионарный симпатический нейрон и его волокно; 5 – преганглионарный симпатический нейрон и его волокно; 6 – преганглионарный парасимпатический нейрон и его волокно; 7 – аксон чувствительного нейрона, передающий восходящие сигналы в ЦНС.
Клетки II типа более крупные, их перикарионы овальной или круглой формы с гладкой поверхностью, при импрегнации серебром имеют темную цитоплазму и светлое ядро с темным ядрышком. От тела клетки отходит до пяти длинных отростков одинакового диаметра. Среди них морфологически трудно различить аксон и дендриты. Отростки, как правило, покидают узел. Клетки II типа являются чувствительными нейронами. Их дендриты образуют разнообразные рецепторные окончания на гладких миоцитах, ганглиях и других элементах. Аксоны образуют синапсы на клетках I, замыкая местную рефлекторную дугу. Кроме того, они отдают коллатерали, оканчивающиеся синапсами на нейронах превертебральных симпатических узлов, через которые чувствительные импульсы от собирательных афферентных нейронов ЖКТ достигают ЦНС.

Рис. 11. Фрагмент вегетативного ганглия МНС. Импрегнация азотнокислым серебром.
1 – клетка Догеля I типа; 2 – её аксон; 3 – клетка Догеля II типа; 4 – ядра глиоцитов; 5 – нервные волокна
Клетки III типа являются местными вставочными нейронами. Их перикарионы имеют овальную или неправильную форму, от них отходят длинный аксон и большое количество коротких дендритов различной длины. Дендриты не выходят за пределы узла, образуют синапсы с клетками II типа. Аксон направляется в другие узлы и вступает в синаптические контакты с клетками I типа.
Клетки III типа встречаются редко, и они мало изучены. Что касается клеток Догеля I и II типов, то они содержатся в значительных количествах в интрамуральных ганглиях всех органов, имеющих метасимпатическую нервную систему.
Исследование интрамуральных нервных аппаратов аллогенно пересаженных сердец 1 — 2 месячных щенков реципиентам того же возраста показало, что через 1 — 5 суток рецепторные окончания и преганглионарные волокна центрального происхождения погибают, а собственные интракардиальные нервные элементы сохраняются и выглядят вполне нормально. Через один месяц большая часть нейронов в узлах представлена дифференцированными мультиполярными клетками. Через 20 – 30 суток появляются рецепторные аппараты, образованные клетками Догеля II типа.
У человека в составе энтеральной нервной системы насчитывается около 108 нейронов – примерно столько же, сколько их содержится в спинном мозге. Разумеется, разнообразие нейронов энтеральной МНС не ограничивается описанными ещё в конце XIX века тремя типами по А.С. Догелю. В настоящее время по совокупности ультраструктурных, иммунохимических, физиологических и других критериев выделены более 10 основных типов нейронов. При этом ассоциативные и эфферентные нейроны могут оказывать на другие нервные или эфферентные (гладкомышечные, секреторные) клетки возбуждающее, тоническое или тормозное действие. Одним из основных видов синаптической передачи в МНС наряду с адренергической и холинергической является также пуринергическая.
К важным морфологическим особенностям узлов энтеральной части МНС, как и остальных вегетативных узлов, относится то, что все без исключения отростки их нейронов представляют собой безмиелиновые проводники (Рис.12), обладающие малыми скоростями передачи нервных импульсов. Интрамуральные метасимпатические ганглии, особенно энтеральные, отличаются от других автономных ганглиев рядом ультраструктурных особенностей. Они окружены тонким слоем глиальных клеток.
Метасимпатическая нервная система человека
Характерная для внеорганных узлов капсула из периневрия и эпиневрия в них отсутствует. Внутри узлов также не содержатся фибробласты и пучки коллагеновых волокон; они встречаются только снаружи от базальной мембраны капсулы из глиоцитов. Перикарионы нервных клеток и их многочисленные отростки заключены в плотный, как в ЦНС, нейропаль. Во многих местах их перикарионы прилежат вплотную друг к другу и не разделены отростками клеток глии.
Межклеточные пространства между нейронами составляют 20 нм. В узлах содержатся многочисленные глиоциты с округлым ядром, богатым гетерохроматином, в их цитоплазме различаются митохондрии, полисомы, другие основные органеллы и пучки глиофиламентов. Кроме того, узлы, как правило, снабжены чувствительными нервными окончаниями. (Рис. 13).

Рис. 12. Ультраструктура безмиелинового нервного волокна. Рисунок с электронограммы с изменениями.
1 – цитоплазма клетки Шванна; 2 – ядро клетки Шванна; 3 – нервные волокна (осевые цилиндры); 4 – оболочка клетки Шванна; 5 – мезаксоны.

Рис. 13. Чувствительные нервные окончания в ганглии кишечного сплетения. Импрегнация по Бильшовскому – Гросс.
Результаты изучения строения и функций метасимпатической нервной системы имеет несомненное практическое значение. Так, к числу распространенных заболеваний желудочно–кишечного тракта относится болезнь Гиршпрунга. У новорожденных она наблюдается с частотой 1: 2000 – 3000, встречается и у взрослых. Причиной болезни является отсутствие и недостаточность развития нервных узлов в межмышечном и подслизистом нервных сплетениях многих сегментов толстой кишки. Эти сегменты кишки спазмированы, а вышележащие резко расширены вследствие нарушения проходимости химуса. Эти проявления болезни Гиршпрунга являются еще одним подтверждением того, что нормальный тонус и двигательная активность кишечника регулируются энтеральной метасимпатической нервной системой. В нетипичных случаях отсутствие узлов (аганглиоз) наблюдается не только в толстой, но и в тощей кишке, желудке и пищеводе, что сопровождается определенными нарушениями функций этих органов. Кроме аганглиоза, при этом заболевании наблюдаются изменения в имеющихся узлах: уменьшение количества нейронов, дистрофические нарушения в их перикарионах, ненормальная извилистость и гиперимпрегнация нервных волокон.
В сердце, как и в ЖКТ, собственная метасимпатическая нервная система имеет решающее значение в регулировании согласованной работы всех элементов органа.
Метасимпатическая нервная системаМНСМногочисленные небольшие скопления нервных клеток, входящие в состав обширных нервных сплетений в стенках внутренних органов (желудочно-кишечного тракта, сердца и др.), иногда относят к парасимпатическому отделу вегетативной нервной системы на том основании, что в морфологических работах легко обнаруживаются синаптические контакты между этими клетками и волокнами блуждающего нерва.
МНС
Интрамуральная нервная система образуется в результате миграции пронейробластов по симпатическим и парасимпатическим нервным стволам.
Вегетативная иннервация сердца: Метасимпатическая интрамуральная нервная системаЛокализация энтеральной нсМетасимпатическая нервная система, интрамуральные нервные сплетения обнаружены в сердце, всех полых органах, но глубже изучены на примере иннервации желудка и кишечника. В этих отделах желудочно-кишечного тракта интрагастральная и энтеральная нервная система представлена настолько обильно, что по числу нейронов (108 единиц) сопоставима со спинным мозгом. Это дает основание для образного названия ее «брюшной мозг».
Физиологические и гистохимические исследования позволяют выявить нейроны, выделяющие в качестве предполагаемых передатчиков
По ответам на длительный толчок деполяризующего тока все эн-теральные нейроны межмышечного сплетения можно разделить на два типа: первый - тип S и второй - тип АН. Нейроны типа S отвечают на это раздражение длительной серией спайков, а нейроны типа АН - лишь одним или двумя спайками, которые сопровождаются сильной и длительной (4-20 с) следовой гиперполяризацией, отсутствующей у типа S. Спайк у нейронов типа S обусловливается натриевой, а у нейронов типа АН - натриевой и кальциевой проводимостью мемб-раны. ПМ - продольная мышца, МС - межмышечное сплетение, К.М - круговая мышца, ПС - подслизистое сплетение, С - слизистая оболочка; указаны нейроны, содержащие или выде-ляющие ацетилхолин [А X}, серотонин (5-гидрокситриптамин (5-ГТ)) и различные пептиды (вызывающие возбуждающие (+) или тормозные МХР - мускариновые холинорецепторы, а-А Р - альфа-адренорецепторы. |
Метасимпатическая нервная система человека
Автономная (вегетативная) нервная система, systema nervo-sutn autonomicum, - часть нервной системы, осуществляющая иннервацию сердца, кровеносных и лимфатических сосудов, внутренностей и других органов. Эта система координирует работу всех внутренних органов, регули-рует обменные, трофические процессы, поддерживает постоянство внутренней среды орга-низма.
Автономная (вегетативная) нервная система подразделяется на центральный и периферический отделы. К центральному отделу относятся: 1) парасимпатические ядра III, VII, IX и X пар черепных нервов, лежащие в мозговом стволе (mesencephalon, ports, medulla oblongala); 2) вегетативное (симпатиче-ское) ядро, образующее боковой промежуточный столб, columna intermediolateralis (autonomica), VIII шейного, всех грудных и двух верхних поясничных сегментов спинного мозга (Cvni, Thi - Lu); 3) крестцовые парасимпатические ядра, nuclei parasym-pathici sacrales, залегающие в сером веществе трех крестцовых сегментов спинного мозга (Sn-Siv).
К периферическому отделу относятся: 1) вегетатив-ные (автономные) нервы, ветви и нервные волокна, па., rr. et neurofibrae autonomici (viscerates), выходящие из головного и спинного мозга; 2) вегетативные (автономные, висцеральные) сплетения, plexus autonomici (viscerates); 3) узлы вегетативных (автономных, висцеральных) сплетений, ganglia plexum autono-micorum (viscerdlium); 4) симпатический ствол, truncus sympathicus (правый и левый), с его узлами, межуз-ловыми и соединительными ветвями и симпатическими нервами; 5) концевые узлы, ganglia termindlia, парасимпатической части вегетативной нервной системы.
Нейроны ядер центрального отдела вегетативной нервной си-стемы являются первыми эфферентными нейронами на пути от ЦНС (спинной и головной мозг) к иннервируемому органу. Нервные волокна, образованные отростками этих нейронов, носят название предузловых (преганглионарных) волокон, так как они идут до узлов периферической части вегетативной нервной системы и заканчиваются синапсами на клетках этих узлов. Вегетативные узлы входят в состав симпатических стволов, круп-ных вегетативных сплетений брюшной полости и таза. Преганглионарные волокна выходят из мозга в соста-ве корешков соответствующих черепных нервов и передних кореш-ков спинномозговых нервов. Узлы периферической части вегетативной нервной системы содержат тела вторых (эффекторных) нейронов, лежащих на пути к иннервируемым органам. Отростки этих вторых нейронов эфферентного пути, несущих нервный им-пульс из вегетативных узлов к рабочим органам, являются послеузелковыми (постганглионарными) нервными волокнами.
В рефлекторной дуге вегетативной части нервной системы эфферентное звено состоит не из одного нейрона, а из двух. В целом простая вегетативная рефлекторная дуга представлена тремя нейронами. Первое звено рефлекторной дуги - это чувствительный нейрон, тело которого располагается в спин-номозговых узлах и в чувствительных узлах черепных нервов. Второе звено рефлекторной дуги является эфферентным, поскольку несет импульсы из спинного или головного мозга к рабочему органу. Этот эфферентный путь вегетативной рефлек-торной дуги представлен двумя нейронами. Первый из этих ней-ронов, второй по счету в простой вегетативной рефлекторной дуге, располагается в вегетативных ядрах ЦНС. Его можно на-зывать вставочным, так как он находится между чувствительным (афферентным) звеном рефлекторной дуги и вторым (эфферент-ным) нейроном эфферентного пути. Эффекторный нейрон пред-ставляет собой третий нейрон вегетативной рефлекторной дуги. Тела эффекторных (третьих) нейронов лежат в периферических узлах вегетативной нервной системы.
Метасимпатическая нервная система — совокупность микроганглионарных образований, находящихся в стенке различных органов, характеризующихся двигательной активностью — метасимпатическая нервная система миокарда, желудочно-кишечного тракта, сосудов, мочевого пузыря, мочеточников. Микроганглии включают 3 вида нейронов: чувствительные, двигательные, вставочные.
Значение метасимпатической нервной системы.
Метасимпатическая нервная система образует местные рефлекторные реакции и включает в себя все компоненты рефлекторных дуг. Благодаря метасимпатической нервной системе внутренние органы могут работать без участия центральной нервной системы. Для изучения метасимпатической нервной системы брали изолированное сердце. В правое предсердие вводили балон с воздухом — растяжение предсердия — приводило к увеличению частоты сердечных сокращений. Внутреннюю поверхность сердца обработали анестетиком и повторили эксперимент — работа сердца не изменялась. Таким образом, внутри сердца есть рефлекторные дуги. Метасимпатическая нервная система обеспечивает передачу возбуждения с эсктраорганной нервной системы на ткань органа — таким образом метасимпатическая нервная система посредник между симпатической нервной системой (парасимпатической нервной системой) и тканью органа. Чаще на метасимпатическую нервную систему образует синапсы парасимпатическая нервная система, чем симпатическая нервная система.
Метасимпатическая нервная система регулирует органный кровоток.
БИЛЕТ № 33
- Локтевой сустав: строение, движения, мышцы, приводящие его в движение. Кровоснабжение, иннервация.
- Наружные женские половые органы. Кровоснабжение, иннервация.
- Вегетативные узлы головы.
Парасимпатичного відділу вегетативної нервової системи
До парасимпатическому відділу вегетативної нервової системи відносяться парасимпатические ядра, утворені парасимпатическими нейронами (центральна частина парасимпатичного відділу вегетативної системи), вузли і парасимпатичні нервові волокна.
Парасимпатический відділ вегетативної нервової системи має такі особливості будови:
1) утворений нервовими волокнами, що відходять від нейронів, розташованих в ядрах середнього мозку (у складі окорухового черепного нерва), довгастого мозку (у складі лицьового, язикоглоткового і блукаючого черепних нервів) і крижового відділу спинного мозку (у складі тазових спинномозкових нервів). Парасимпатичні волокна, що виходять з головного і спинного мозку, йдуть до нервових вузлів;
2) нервові вузли лежать близько органу або в иннервируемой органі (входять до складу вегетативних сплетінь);
3) прегангліонарних волокна довгі, так як йдуть від ЦНС до органу;
4) постгангліонарні волокно коротке, так як знаходиться безпосередньо в органі.
Функції парасимпатичної іннервації. Парасимпатична нервова система іннервує м’язи очей, мови, слинні залози, трахею і бронхи, легені, всі органи травлення, серце, нирки, сечовий міхур, сечоводи і інші внутрішні органи, а також деякі кровоносні судини. Передача імпульсів з постгангліонарних волокна на орган здійснюється за допомогою медіатора ацетилхоліну.
Велика частина порожніх внутрішніх органів (серце, бронхи, сечовий міхур, травний тракт, матка, жовчний міхур,
жовчні шляхи і ін.) поряд з симпатичної і парасимпатичної іннервації мають власний місцевий механізм регуляторноїлції — метасимпатична нервову систему.
Місцем локалізації метасімпатіческой нервової системи є інтрамуральні ганглії, що залягають в товщі стінок порожнистих органів і ізольовані від навколишніх тканин спеціальними бар’єрами.
Метасимпатична нервова система має власний чутливий нейрон, інтернейрон, еффекторний нейрон і медіаторна ланка. Тіла нейронів метасімпатіческой нервової системи несуть безліч синапсів, а в відростках цих нейронів міститься велика кількість бульбашок з медіаторами. Метасимпатична нервова система іннервує тільки внутрішні органи.
Функції метасімпатіческой нервової системи. Метасимпатична нервова система програмує і координує рухову, секреторну і всмоктувальної активність органів, активність місцевих ендокринних елементів і локальний кровотік. Вона визначає здатність органів ритмічно скорочуватися з певною частотою і амплітудою без впливу ззовні під впливом метаболічних змін в самому органі.
Передача збудження в нейронах, що становлять ганглії метасімпатіческой системи, здійснюється за участю ацетилхоліну і норадреналіну.
В синапсах постгангліонарних волокон виділяються різноманітні речовини — ацетилхолін, норадреналін, АТФ, аденозин і ін.
